Archivio Micologico
-
Numero contenuti
14587 -
Iscritto
-
Ultima visita
Tipo di contenuto
Profili
Forum
Orchidee
Diventa Socio
Calendario
Tutti i contenuti di Archivio Micologico
-

Hortiboletus bubalinus (Oolbekk. & Duin) L. Albert & Dima, 2015
Archivio Micologico ha postato un topic nell'area Funghi Commestibili
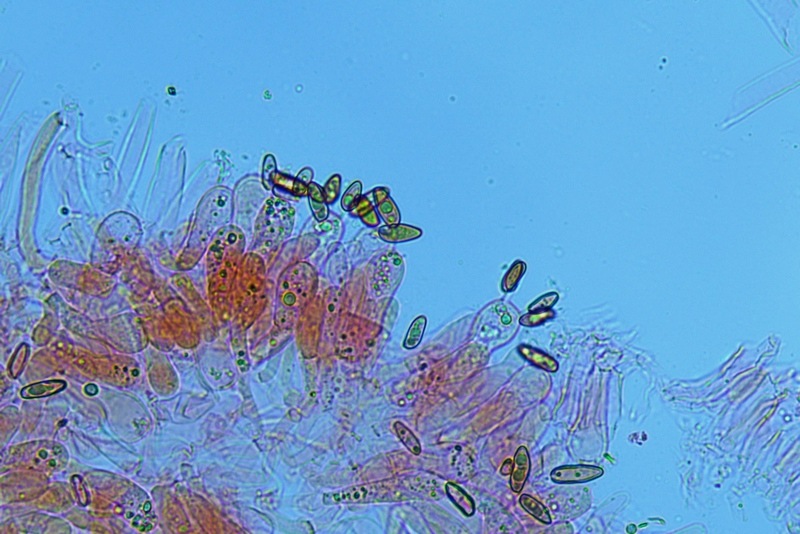
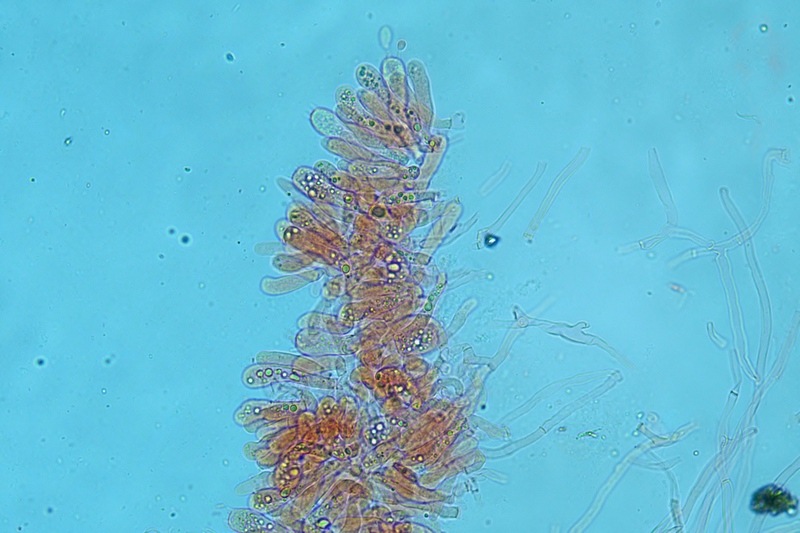
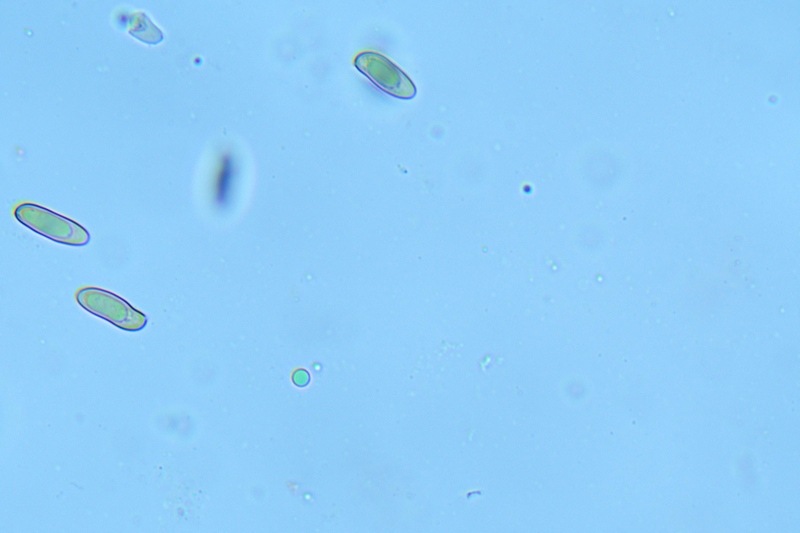
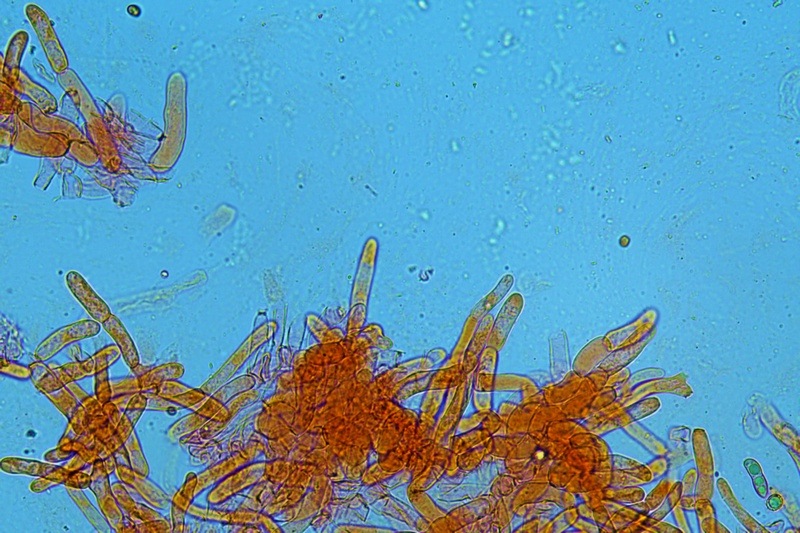
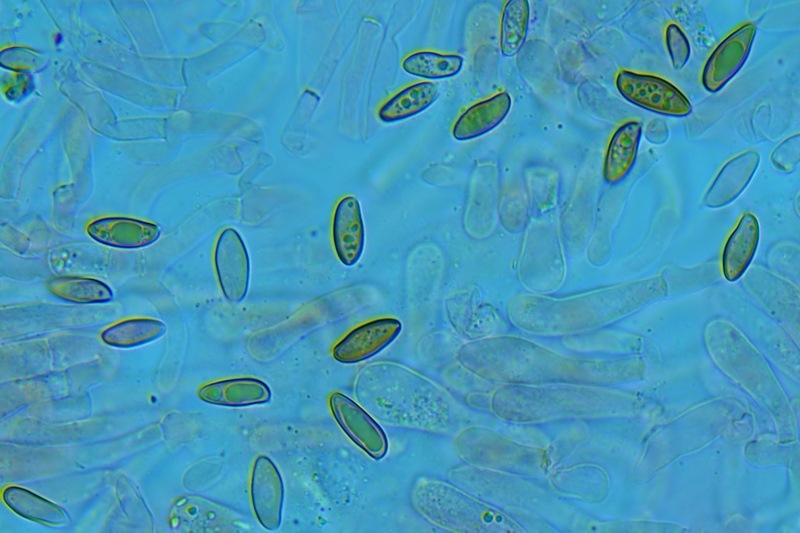
Hortiboletus bubalinus (Oolbekk. & Duin) L. Albert & Dima, 2015 Tassonomia Divisione Basidiomycota Classe Agricomycetes Ordine Boletales Famiglia Boletaceae Sinonimi Xerocomellus bubalinus (Oolbekk. & Duin) Mikšík 2014 Xerocomus bubalinus (Oolbekk. & Duin) Redeuilh 1993 Boletus bubalinus Oolbekk. & Duin 1991 Etimologia L'epiteto Hortiboletus deriva dal latino hortus = giardino e da boletus che avrebbe una etimologia non chiara, il termine potrebbe derivare dal greco βωλήτης [bolétes] = una sorta di fungo, oppure da βῶλος [bólos] = zolla, terra, in riferimento al substrato di crescita. Ci sono comunque opinioni diverse che ritengono che βῶλος debba intendersi con il significato di sfera, poiché il giovane cappello in questo genere assume sempre una forma globosa. Per altri ancora deriverebbe dal latino bolites, definizione con la quale gli antichi romani indicavano i pregiati funghi commestibili, all'esordio per citare i soli ovoli, oggi Amanita caesarea, ma in seguito utilizzato in modo esteso anche per indicare i porcini. L'epiteto bubalinus deriva dal latino būbălus = bufalo, per indicare il colore del mantello del bufalo. Cappello Inizialmente emisferico, poi espanso e appianato, a maturità lievemente depresso al centro; la cuticola si presenta vellutata di colore nocciola e si decolora verso il margine assumendo tipiche tonalità rosate. Spesso nei soggetti adulti risulta screpolata e fessurata al margine, leggermente eccedente nei soggetti giovani. Pori e tubuli Nei soggetti giovani i pori sono piccoli, rotondeggianti e irregolari, di colore giallo, adnati al gambo o brevemente decorrenti, a maturazione sono ampi, angolosi, olivastri, verdastri. Tubuli concolori ai pori. Gambo Cilindrico, spesso anche leggermente clavato verso la base, giallastro con fiammature rossastre per tre quarti, bianco alla base dove si evidenziano residui miceliali giallastri. Carne La carne del cappello alla sezione presenta una colorazione rosa, innata, non da viraggio. In sezione longitudinale, oltre alla carne del cappello naturalmente rosa, si apprezzano diversi viraggi: nella parte superiore del gambo azzurro; nella parte mediana verdastro; alla base brunastro. Il micelio al piede è giallastro. L'odore può essere definito come gradevole e fruttato, di sapore dolce e fungino. Habitat Tigli e Pioppi di varie specie, privilegiando giardini e parchi, da cui il nome del genere. Commestibilità e tossicità Commestibile eliminando il gambo perché fibroso, coriaceo e quindi poco digeribile. Microscopia Cheilocistidi non molto frequenti, fusiformi ma anche cilindrici a volte con apice incurvato, settati alla base. Pleurocistidi rari simili ai cheilocistidi. GAF non osservati. Pileipellis una palizzata di ife cilindriche, catenulate e brevemente settate, a parete incrostata, con pigmento ocraceo a placche, ife terminali con apice ristretto e arrotondato. Basidi clavati, bisporici e tetrasporici, con guttule oleose giallastre. Spore (9,8) 10,2-13,7 (14,6) × (3,5) 4,1-5,0 (5,4) µm; Q = (2,2) 2,4-3,0 (3,2); N = 44; Media = 12,0 × 4,5 µm; Qm = 2,7; lungamente ellittiche, subfusiformi, mono- e pluri-guttulate con pigmento giallastro, con evidente depressione sopra l'apicolo. Stipitipellis costituita da cellule allungate, a textura intricata. Caulocistidi e Caulobasidi presenti sulla parte alta del gambo. Specie simili Hortiboletus rubellus (Krombh.) Simonini, Vizzini & Gelardi, che presenta simili caratteri microscopici ed analoghe esigenze ecologiche, si differenzia per le colorazioni decisamente rossastre del pileo e della base dello stipite, per la presenza delle classiche punteggiature rosse nella carne alla base del gambo; per l'assenza del colore rosa innato nella carne del cappello che diversamente è gialla, lievemente rosa solo sotto la cuticola, inoltre, non presenta colori brunastri nella carne alla base dello stipite, ma colorazioni rosso arancio. Hortiboletus engelii (Hlaváček) Biketova & Wasser, presenta colorazioni analoghe del pileo e della corteccia del gambo, ma presenta tipiche punteggiature rosse nella carne alla base del gambo; non mostra il colore rosa innato nella carne del cappello che diversamente è gialla, non presenta colori brunastri nella carne alla base dello stipite che risulta essere di una tonalità giallo pallido. Xerocomellus chrysenteron (Bull.) Šutara, presenta una ecologia completamente diversa, strettamente legata alle fresche foreste di conifere e alle faggete di altura. Osservazioni Nella monografia Xerocomus s.l. Fungi Europæi pubblicata nel 2003, si legge che all'epoca si riteneva questa specie fosse presente e conosciuta solo nel territorio dell'Olanda, dove fruttificava da agosto a settembre in aree urbane associate a Tilia e Popolus alba, ad oggi tale convinzione ecologica è stata aggiornata con nuovi dati, infatti Hortiboletus bubalinus risulta presente in quasi tutto il territorio italiano, in particolare nei giardini e parchi planiziali, con habitat tipicamente mediterraneo. Note nomenclaturali Dal 2015 la specie è stata segregata nel neogenere Hortiboletus grazie ad indagini molecolari di Gelardi et al.; tale genere annovera anche Hortiboletus rubellus (Krombh.) Simonini, Vizzini & Gelardi, Hortiboletus campestris (A.H. Sm. & Thiers) Biketova & Wasser e Hortiboletus engelii (Hlaváček) Biketova & Wasser. Bibliografia LADURNER, H. & SIMONINI, G., 2003. Xerocomus s.l. Fungi Europæi. Vol. 8. Alassio (SV): Ed. Candusso. ŠUTARA, J., 2008. Xerocomus s. l. in the light of the present state of knowledge. Czech Mycol. 60(1): 29-62. [Data di accesso: 10/04/2022]. Scheda di proprietà AMINT realizzata da Pietro Curti, e Giovanni Galeotti - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Umbria; Dicembre 2021; Foto e microscopia di Giovanni Galeotti. (exsiccatum GG211222-01) Soggetti rinvenuti nel giardino di una villa privata, l'habitat dei ritrovamenti comprende tiglio, pioppo, leccio e cedro. La carne del pileo alla sezione, evidenzia una bella colorazione rosa innata. Cheilocistidi. Osservazione in rosso Congo, a 400×. Cheilocistidi. Osservazione in rosso Congo, a 1000×. Basidi. Osservazione in rosso Congo. Basidi. Osservazione in rosso Congo, a 400×. Basidi. Osservazione in rosso Congo, a 1000×. Pileipellis. Osservazione in rosso Congo, a 400×. Pileipellis. Osservazione in rosso Congo, a 1000×. Pileipellis. Osservazione in acqua, a 400×. Pileipellis. Osservazione in acqua, a 1000×. Spore(9,8) 10,2-13,7 (14,6) × (3,5) 4,1-5,0 (5,4) µm; Q = (2,2) 2,4-3,0 (3,2); N = 44; Media = 12,0 × 4,5 µm; Qm = 2,7. Osservazione in acqua, a 1000×.


.jpg.988f1004e3ff21ea732e79f4ff21e36c.jpg)
.jpg.fa4bb9716989d1bb88d057b8b48080b6.jpg)
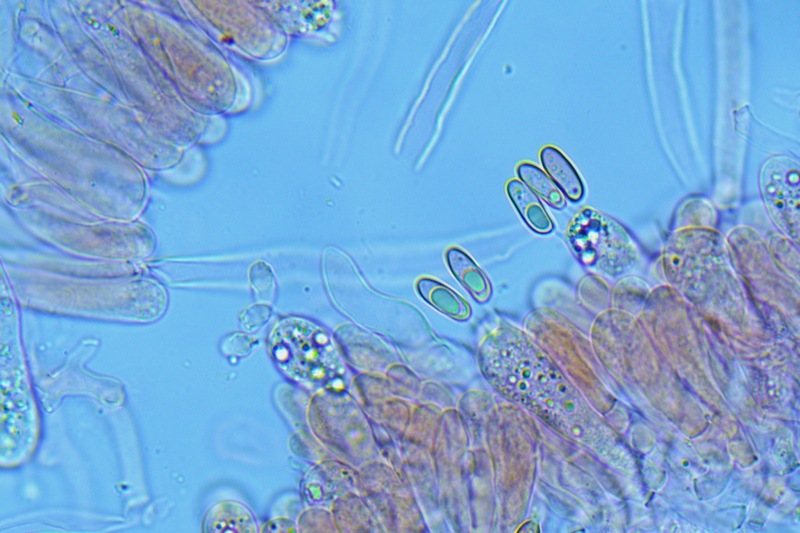
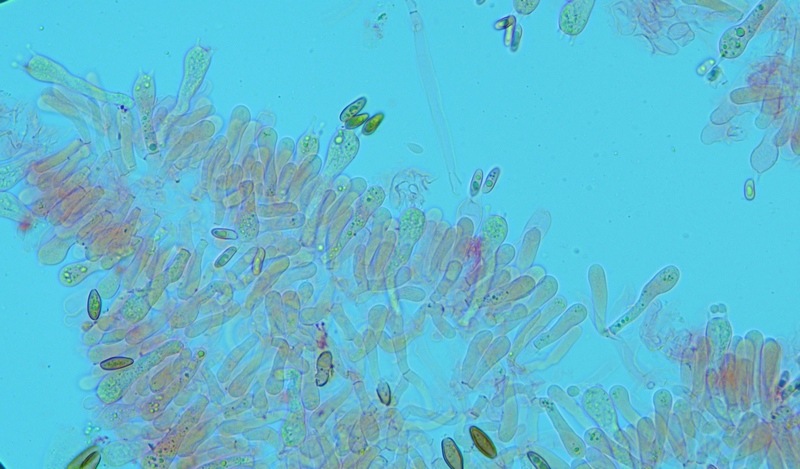
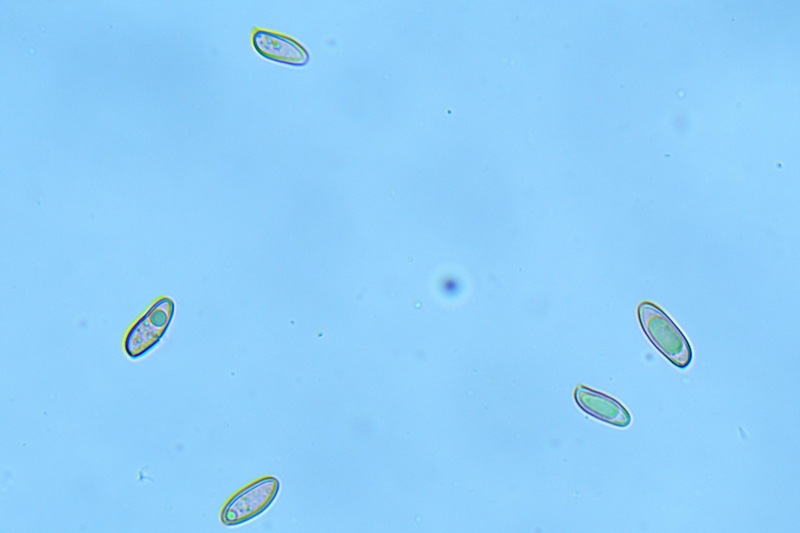
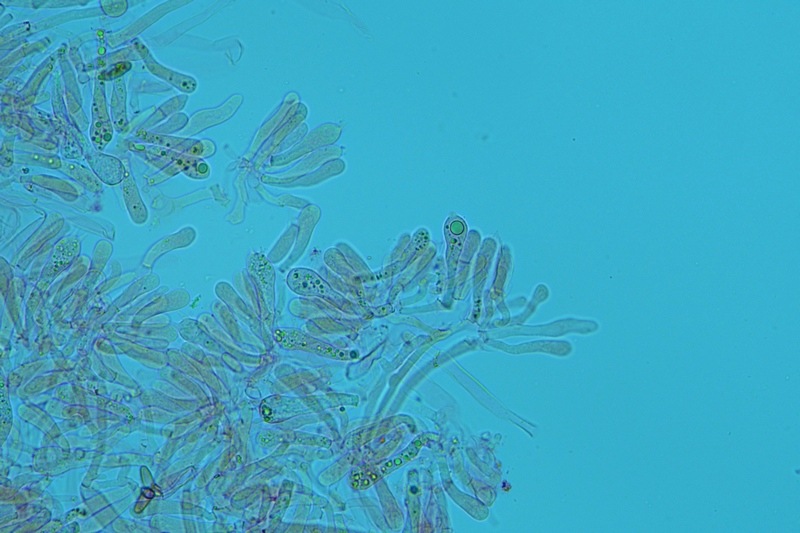
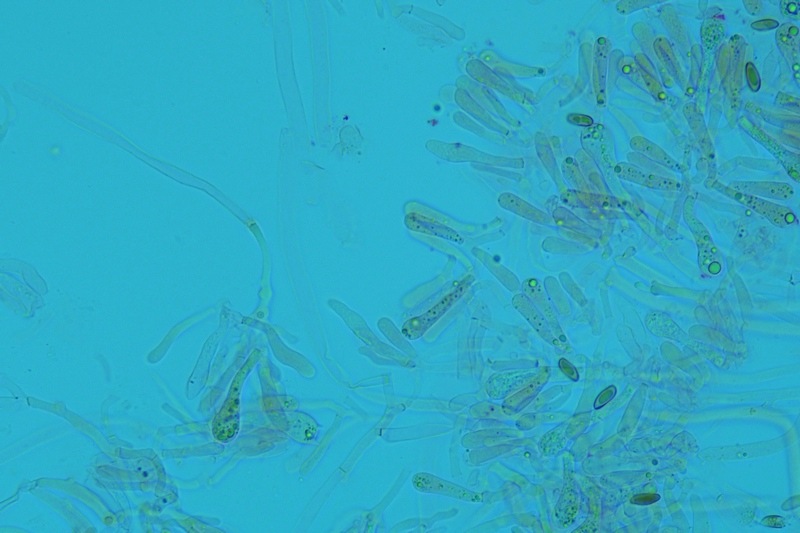
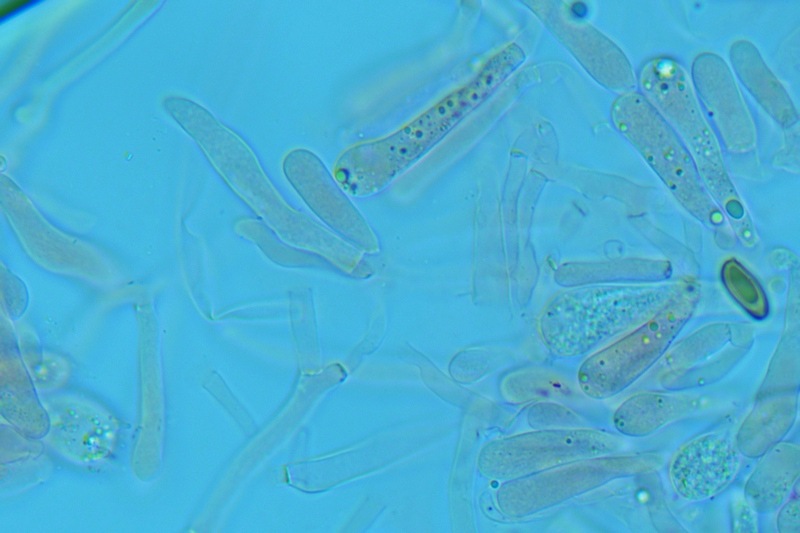
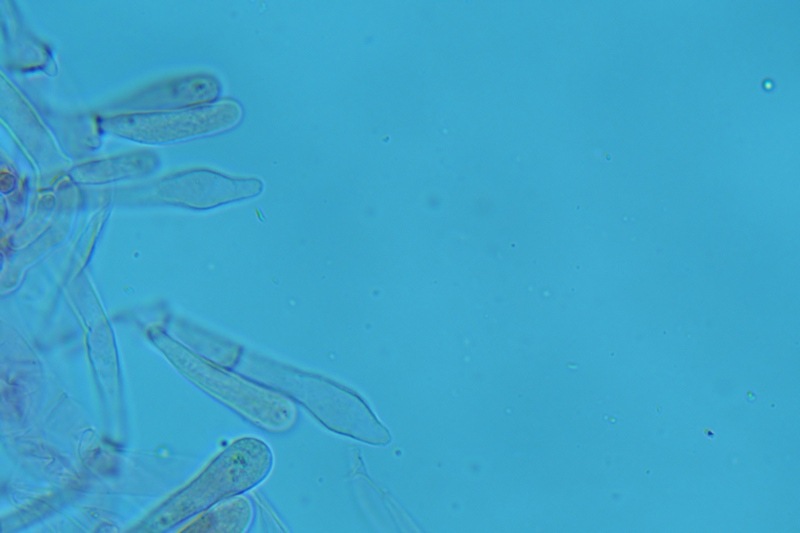
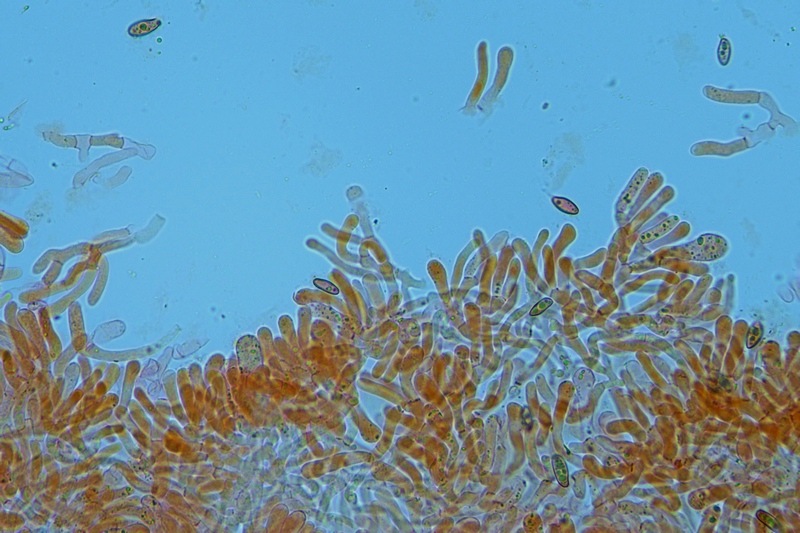
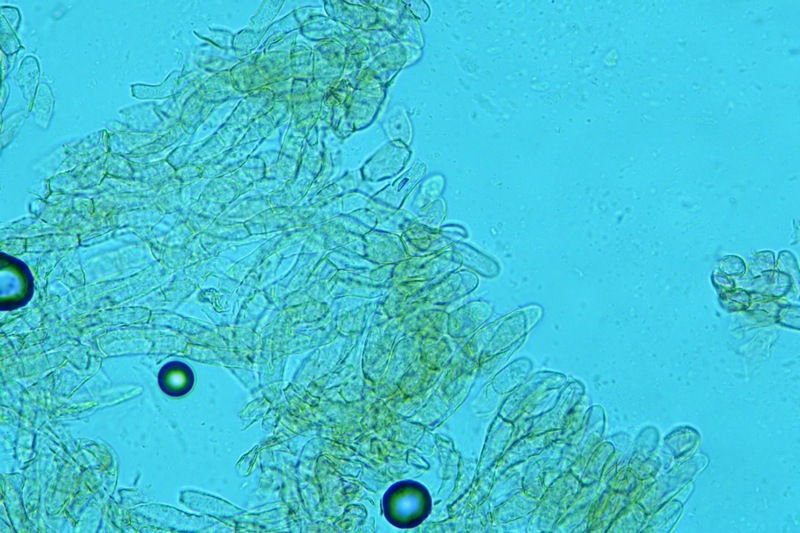
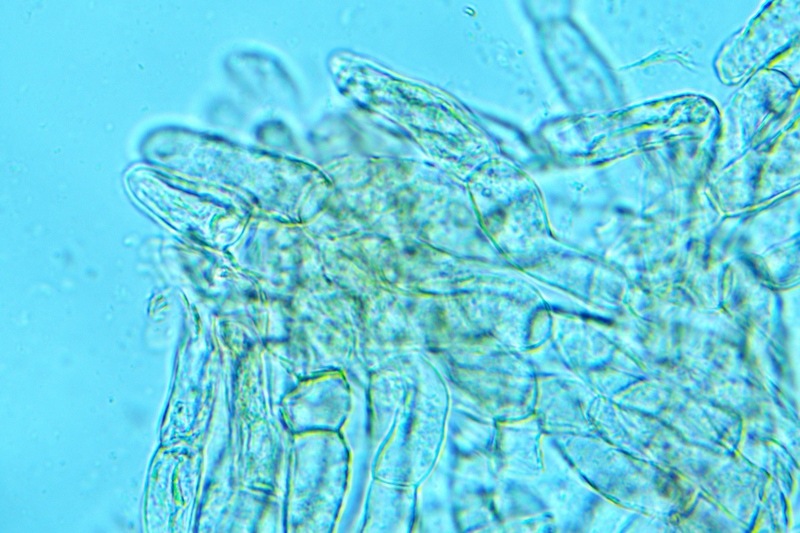
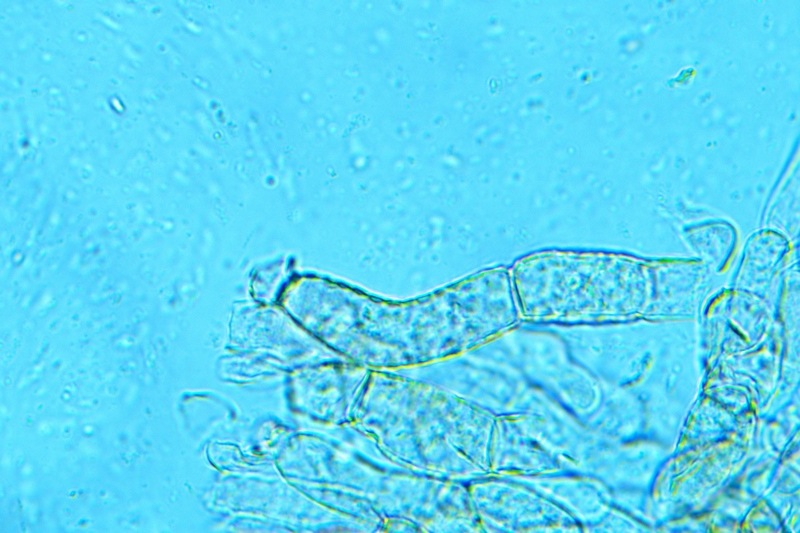
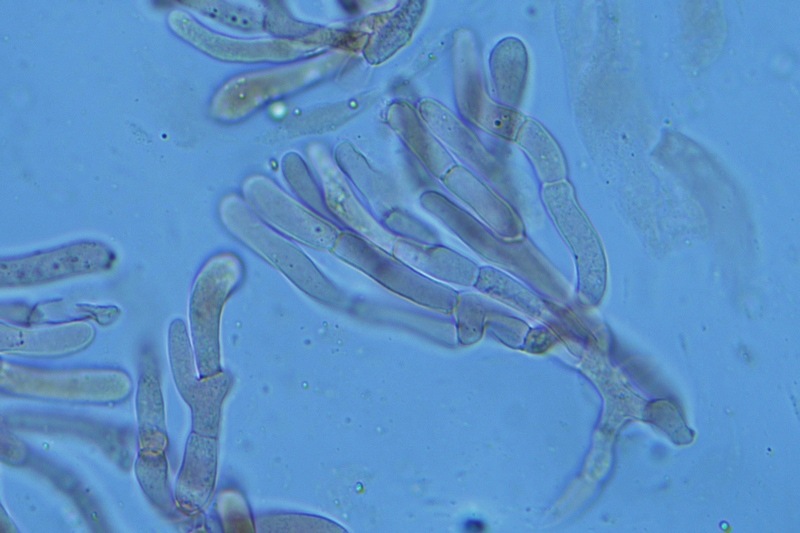
-
Inocybe dulcamara (Pers.) P. Kumm. 1871
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Velenosi
Inocybe dulcamara (Pers.) P. Kumm.; Regione Lazio; Marzo 2007; Foto di Felice di Palma. Ritrovamento in bosco misto con Pino e Leccio, odore gradevole, mielato.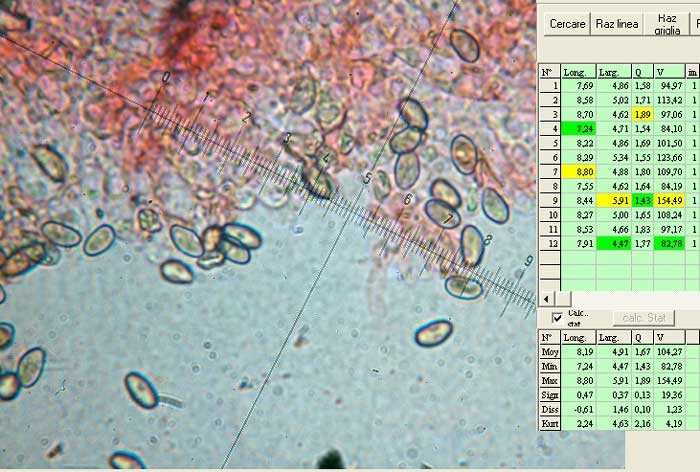
-
Phlebia radiata Fr. : Fr. 1821
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Phlebia radiata Fr. : Fr. 1821 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Polyporales Famiglia Meruliaceae Descrizione Fungo di colore arancio acceso, con imenoforo a crescita radiale. Regione Toscana; Ottobre 2021; Foto e descrizione di Stefano Rocchi. -
Hericium alpestre Pers. 1825
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Hericium alpestre Pers. 1825 Tassonomia Divisione Basidiomicota Classe Agaricomycetes Ordine Russulales Famiglia Hericiaceae Foto e Descrizioni Su tronco a terra di Abete bianco. Si caratterizza per le spine non ramificate, lunghe fino a 2 cm, raggruppate in ciuffi lungo le diramazioni del basidioma. Microscopia Spore 5,8-6,3 × 5-6 µm, subglobose, finemente verrucose e amiloidi. Specie simili Hericium cirrhatum (Pers. : Fr.) Nikol. si distingue per il basidioma pileato e le spore più piccole. Hericium erinaceus (Bull. : Fr.) si differenzia per le spine lunghe 4-5 cm. Hericium coralloides (Scop. : Fr.) Pers. si distingue per le spine più corte, ramificate, disposte nella parte inferiore delle diramazioni del basidioma e per le spore più piccole. Regione Toscana; Ottobre 2021; Foto, descrizione e microscopia di Stefano Rocchi. Spore 5,8-6,3 × 5-6 µm, subglobose, finemente verrucose e amiloidi. Osservazione in Melzer, a 1000×. -
Arrhenia monsducalis Albanese, Battistin, Berna, Boragine, Ercole & Vizzini 2021 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Hygrophoraceae Etimologia L'epiteto Arrhenia è dedicato al micologo Johan Peter Arrhenius (1811-1889). L'epiteto monsducalis si riferisce a via Monte Ducale, Minturno (LT) Lazio, Italia; luogo di raccolta dell’olotipo. Basidioma Pileato-stipitato, lamellato, omogeneo, piuttosto slanciato, scarno e minuto; terricolo. Cappello 10-19 mm di diametro, convesso, con zona discale profondamente depressa, omfaloide-imbutiforme; margine striato per trasparenza, talvolta sinuoso e ondulato e leggermente involuto; cuticola igrofana, liscia, con colori variabili da bruno a grigio-bruno, con attenuazione dell’intensità cromatica verso il margine. Lamelle Decorrenti, subconcolori al pileo, ma più pallide, con riflessi bluastri, talvolta cangianti al viola, subrade, intercalate da numerose lamellule di secondo e terzo ordine; margine concolore, intero. Gambo 10-16 mm × 2-3,5 mm, pieno, centrale, esile, cilindrico, svasato all’inserzione con il pileo, attenuato in basso; solcato, liscio, di colorazione sostanzialmente uguale a quella delle lamelle, grigio chiaro nella parte alta, tende al grigio marrone verso la base; presenza di micelio basale di colore bianco. Carne Esigua, elastica, di colore bianco sporco, odore molto pronunciato di pelargonio, sapore non testato. Habitat e modo di crescita Gregario, in microambiente umicolo-muscicolo in macchia mediterranea, con presenza di Quercus suber L., Q. virgiliana (Ten.) Ten., Erica arborea L. e Myrtus communis L. Fenologia Novembre. Microscopia Spore (5,7) 5,9 - 7,2 (7,8) × (3) 3,2 - 3,9 (4,2) µm Q = (1,5) 1,6 - 2 (2,4); N = 30; Media = 6,5 x 3,5 µm; Qm = 1,9; bianche in massa, da ellissoidali ad ovoidali, lisce, ialine, con piccole granulazioni interne, prive di poro germinativo e con apicolo evidente. Basidi (21,1) 28,6 - 36 (42,6) × (4,6) 5,1 - 6,8 (8,6) µm; N = 30; Media = 33.1 × 6 µm; cilindrico-claviformi, tetrasporici o bisporici, provvisti di sterigmi lunghi 3,97 - 5,9 µm. Basidioli simili ai basidi (19,9) 25,8 - 38,1 (41,1) × (3,3) 3,7 - 6 (6,9) µm; N = 30 Media = 31,6 × 4,9 µm. Trama lamellare da subregolare a irregolare, costituita da ife intrecciate larghe 1,5 - 3,5 µm. Cistidi di ogni tipo non osservati. Pileipellis costituita da un ixocutis ife cilindriche parallele, settate e immerse in una matrice gelatinosa, larghe 2-6 µm, con granulazioni extracellulari parietali, incrostanti, di colore brunastro. Caulipellis composta da lunghe ife ascendenti 2,3 - 3,2 µm, ialine, parallele, cilindriche e settate, di cui alcune più spesse, che simulano dei caulocistidi, apice arrotondato. GAF assenti in tutte le parti del basidioma. Specie simili Arrhenia velutipes (P.D. Orton) Redhead, Lutzoni, Moncalvo & Vilgalys 2002, si distingue per la presenza dei giunti a fibbia su tutto il basidioma e per la crescita montana. Note tassonomiche Studi molecolari comprendenti i taxa omphalinoidi (Moncalvo et al. 2002, Redhead et al. 2002 e Lodge et al. 2014) hanno progressivamente permesso di meglio delineare la delimitazione morfologica dei vari generi implicati e di chiarire le loro reali affinità filogenetiche. Tali taxa, tutti riferiti a morfologie fungine molto elementari, si sono rivelati appartenere a varie linee evolutive. Il genere Arrhenia, introdotto da Fries (1849), è uno di tali taxa. A lungo considerato un piccolo genere di funghi pleurotoidi con imenoforo ridotto, prevalentemente associati a muschi, da un punto di vista sistematico, è stato dapprima incluso nelle Pleurotaceae (Kühner & Lamoure 1972) e poi nelle Tricholomataceae (Kühner 1980). Più recentemente (Lodge et al. 2014), sulla base di affinità filogenetiche, lo hanno invece incluso nella famiglia Hygrophoraceae, subfam. Hygrophoroideae, tribù Arrheniae, di cui è il genere tipo. Di quest’ultimo raggruppamento fanno parte anche i generi Acantholichen, Cora, Corella, Cyphellostereum, Dyctionema e Eonema. Arrhenia comprende funghi con ampia distribuzione a livello planetario (Redhead 1984, Bon 1987, Barrasa & Rico 2004, Cripps & Horak 2006, Blanco-Dios 2019, Voitk et al. 2020), anche se quella prevalente è nelle aree temperate della regione paleartica, con ambiente di crescita per lo più muscicolo. I basidiomi sono di tipo pileato-stipitato, con contesto sottile e dalle dimensioni piccole, ragion per cui le specie ad esso ascritte hanno silhouette per lo più cifelloide, anche se non mancano taxa ad aspetto pleurotoide o omphalinoide. I colori sono generalmente poco appariscenti, grigio-brunastri, le cuticole opache, a volte squamulose, con margine (corona marginale) spesso da sinuoso ad ondulato; l’imenoforo, del tipo lamellato, ha spesso una densità ridotta, con lamelle da poco fitte a rade, e produce spore bianche in massa, a reazione non amiloide in Melzer; lo stipite, infine, può essere sia corto, sia slanciato. L’indagine microscopica mostra una pileipellis con pigmenti incrostanti ed una trama lamellare spesso irregolare, mentre le ife possono presentare o meno giunti a fibbia; rilevante è anche l’assenza di cistidi. Bibliografia ALBANESE, A., BATTISTIN, E., BERNA, C., BORAGINE, M., ERCOLE, E. & VIZZINI, A., 2021. Nomenclatural Novelties. RMR Bolletino AMER, numero speciale (fuori serie): 34-35. BARRASA, J.M. & RICO, V.J., 2003. The non-omphalinoid species of Arrhenia in the Iberian Peninsula. Mycologia 95(4): 700-713. BARRASA, J.M., RICO, V.J. & VILLAREAL, M., 2003. Arrhenia eburnea, sp. nov. from Spain. Mycotaxon 88: 113-118. BON, M., 1997. Flore mycologique d’Europe 4. CORRIOL, G., 2016. Arrhenia subglobisemen, un nouveau nom pour Agaricus tremulus sensu Persoon, Fries. Bulletin Trimestrel Féderation Mycologique Dauphiné-Savoie 222: 5-20. ELBORNE, S.A., 2008. Arrhenia Fr. Funga Nordica. 1st ed. Eds. H. Knudsen, J. Vesterholt. Copenhagen: Nordsvamp: 226-234. FAVRE, J., 1939. Champignons rares ou peu connus des hauts-marais jurassiens. Bulletin de la Société Mycologique de France 55: 196-219. FAVRE, J., 1960. Catalogue descriptif des champignons supérieurs de la zone subalpine du Parc National Suisse. Ergebnisse der Wissenschaftlichen Untersuchungen des Scheizerischen Nationalparks 6: 323-610 + 8 pl. CLEMENÇON, H., 2012. Cytology and Plectology of the Hymenomycetes. HERTZOG, P., 2005. Notes sur quelques espèces pleurotoïdes. Bulletin Societé Mycologique du Haut Rhin 22: 18-20. KÜHNER, R. & LAMOURE, D., 1972. Agaricales de la zone alpine. Pleurotacées. Le Botaniste 55: 7-37. MONCALVO, J.M., LUTZONI, F.M., REHNER, S.A., JOHNSON, J. & VILGALYS, R., 2000. Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Systematic Biology 49: 278-305. MONCALVO, J.M., VILGALYS, R., REDHEAD, S.A., JOHNSON, J.E. et al. 2002. One hundred and seventeen clades of euagarics. Molecular Phylogenetics Evolution 23: 357-400. NORVELL, L.L., REDHEAD, S.A. & AMMIRATI, J.F., 1994. Omphalina s.l. in North America 1-2. 1: Omphalina wynneae and the genus Chrysomphalina. 2: Omphalina s. Bigelow. Mycotaxon 50: 379-407. POMERLEAU, R., 1980. Flore des champignons au Québec. Montréal, La Presse. REDHEAD, S.A., 1984. Arrhenia and Rimbachia, expanded generic concepts, and a reevaluation of Leptoglossum with emphasis on muscicolous North American taxa. Canadian Journal of Botany 62(5): 865-892. REDHEAD, S.A., LUTZONI, F., MONCALVO, J.M. & VILGALYS, R., 2002. Phylogeny of agarics: partial systematics solutions for core omphaloid genera in the Agaricales (euagarics). Mycotaxon 83: 19-57. SENN-IRLET, B., 1986. Ökologie, Soziologie und Taxonomie alpiner Makromyzeten (Agaricales, Basidiomycetes) der Schweizer Alpen. Dissertation Universität Bern. Manuskript, 252 S. und 39 Taf. VOITK, A., 2017. Arrhenia subglobisemen. Newsletter of Foray Newfoundland and Labrador. Vol VIII (5): 19. Scheda di proprietà AMINT realizzata da Antonio Albanese, Claudio Berna e Marcello Boragine - revisionata da Tomaso Lezzi, approvata dal CLR Micologico di AMINT. Regione Lazio; Novembre 2021; Foto e microscopia di Antonio Albanese e Marcello Boragine. Spore. Osservazione in acqua, a 1000×. Spore. Osservazione in rosso Congo, a 630×. Basidi e basidioli. Osservazione in rosso Congo, a 1000×. Imenio. Osservazione in rosso Congo, a 400×. Pileipellis. Osservazione in acqua, a 630×. Trama lamellare. Osservazione in rosso Congo, a 400×. Caulipellis. Osservazione in rosso Congo, a 400×. Ife della caulipellis, con apice arrotondato. Osservazione in rosso Congo, a 400×.
-
Phyllotopsis nidulans (Pers. : Fr.) Sing. 1936
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Phyllotopsis nidulans (Pers. : Fr.) Sing.; Regione Toscana; Febbraio 2022; Foto di Alessandro Francolini. -
Hygrophorus marzuolus (Fr. : Fr.) Bres. 1893
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Hygrophorus marzuolus (Fr. : Fr.) Bres.; Regione Toscana; Febbraio 2022; Foto di Alessandro Francolini. -
Tremella mesenterica Retz. : Fr. 1769
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Tremella mesenterica Retz. : Fr.; Regione Toscana; Febbraio 2022; Foto di Alessandro Francolini. Minuscoli esemplari, larghi al massimo 2 o 3 millimetri. Esemplari già più sviluppati. -
Bjerkandera adusta (Willd. : Fr.) P. Karst. 1879
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Bjerkandera adusta (Willd. : Fr.) P. Karst.; Regione Toscana; Gennaio 2022; Foto di Alessandro Francolini. Confronto tra le due superfici, la superiore sterile, l' inferiore poroide. La superficie poroide che scurisce al minimo sfregamento. -
Fomitopsis pinicola (Sw. : Fr.) P. Karst. 1881
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Fomitopsis pinicola (Swartz : Fr.) P. Karst.; Regione Toscana; Gennaio 2022; Foto di Alessandro Francolini. Giovani esemplari nati su un pino tagliato. Esemplare più grande. Imenoforo -
Cordyceps militaris (L. : Fr.) Fr. 1818
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Cordyceps militaris (L. : Fr.) Fr.; Regione Toscana; Dicembre 2021; Foto di Alessandro Francolini. Ascomicete che si sviluppa sotto forma di fruttificazione complessiva di periteci; come in altre specie simili i periteci non nascono isolati ma formano delle colonie addensandosi l’un l’altro. I periteci sono immersi nello stroma, un tessuto compatto di ife, lasciando apparire in superficie l’ostiolo, l'apertura che li collega all'esterno. Di solito la presenza degli ostioli è rivelata dalla granulosità della testa stromatica. Teste stromatiche di colore arancione vivo e dalla forma irregolarmente clavata; il gambo sottostante è cilindrico, liscio, non porta nessun peritecio ed è di colore più chiaro rispetto alla testa. La specie fruttifica parassitando larve o crisalidi di lepidotteri interrati. Estratti alcuni esemplari e buona parte del terriccio sottostante, attaccati ai fragili e bianchi cordoncini miceliari, sono state riscontrate solo piccole formazioni globose che si sono disfatte in materiale terroso appena manipolate. Testa stromatica e sue dimensioni. È evidente la presenza di numerosissimi periteci. In sezione si nota lo stroma, con i periteci in esso inseriti. La testa stromatica si mostra vuota all'interno. -
Flammulina velutipes (Curtis : Fr.) Singer 1951
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Flammulina velutipes (Curtis : Fr.) Singer; Regione Toscana; Dicembre 2021; Foto di Alessandro Francolini. Esemplari quasi a fine ciclo. I cappelli hanno perduto gran parte del loro bel colore brillante. In questo cespo i gambi si sono mostrati "vellutati" non solo alla base ma su quasi tutta la superficie: -
Tricholoma caligatum (Viviani) Ricken 1915
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Tricholoma caligatum (Viviani) Ricken; Regione Toscana; Novembre 2021; Foto di Alessandro Francolini. -
Phaeotremella foliacea (Pers.) Wedin, J.C. Zamora & Millanes 2016
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Phaeotremella foliacea (Pers.) Wedin, J.C. Zamora & Millanes; Regione Toscana, Novembre 2021; Foto di Alessandro Francolini. -
Suillus bellinii (Inzenga) Kuntze 1898
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Suillus bellinii (Inzenga) Kuntze; Regione Toscana; Novembre 2021; Foto di Alessandro Francolini. -
Lactarius sanguifluus (Paulet) Fr. 1838
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Lactarius sanguifluus (Paulet) Fr.; Regione Toscana; Novembre 2021; Foto di Alessandro Francolini. -
Auricularia mesenterica (Dicks. : Fr.) Pers. 1822
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Auricularia mesenterica (Dicks. : Fr.) Pers.; Regione Toscana; Novembre 2021; Foto di Alessandro Francolini. Imenoforo con costolature evidenti. Alla sezione la carne si presenta spessa e gelatinosa. -
Hypholoma fasciculare (Huds. : Fr.) P. Kumm. 1871
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Velenosi
Hypholoma fasciculare (Huds. : Fr.) Kumm.; Regione Toscana; Novembre 2021; Foto di Alessandro Francolini. Nascosti sotto un tronco marcescente e caduto a terra. Evidente l'affetto del geotropismo: la nascita al di sotto del tronco costringe i gambi a dirigersi inizialmente in orizzontale o addirittura verso il basso. Ma poi, con lo sviluppo, i gambi si piegano verso l'alto per portare l'imenoforo nelle condizioni ideali per la dispersione delle spore. -
Trichaptum abietinum (Pers. ex J.F. Gmel.) Ryvarden 1972
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Trichaptum abietinum (Pers. ex J.F. Gmel.) Ryvarden; Regione Toscana; Ottobre 2021; Foto di Stefano Rocchi. Su tronco di Abete bianco a terra in una sua forma resupinata. -
Marasmius oreades (Bolton) Fr. 1836
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Marasmius oreades (Bolt. : Fr.) Fr.; Regione Toscana; Novembre 2021; Foto di Alessandro Francolini. Una "saetta" e due "cerchi delle streghe" nei prati. -
Gymnopus fusipes (Bull. : Fr.) Gray 1821
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Gymnopus fusipes (Bull. : Fr.) Gray; Regione Toscana; Novembre 2021; Foto di Alessandro Francolini. -
Hypholoma capnoides (Fr. : Fr.) P. Kumm. 1871
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Hypholoma capnoides (Fr. : Fr.) P. Kumm.; Regione Toscana; Ottobre 2021; Foto e microscopia di Stefano Rocchi. -
Infundibulicybe geotropa (Bull. : Fr.) Harmaja 2003
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Infundibulicybe geotropa (Bull. : Fr.) Harmaja; Regione Toscana; Ottobre 2021; Foto di Stefano Rocchi. -
Lepiota kuehneri Huijsman ex Hora. 1943
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Velenosi
Lepiota kuehneri Huijsman ex Hora 1960; Regione Toscana; Ottobre 2021; Foto di Tomaso Lezzi. Lepiota appartenente al gruppo delle ovispore con cuticola trichodermica, con sub-couche di ife corte di varia lunghezza. Di aspetto macroscopico vicino a Lepiota castanea e a Lepiota cristata, che però appartengono al gruppo delle stenospore cioè delle Lepiota con spore a forma di proiettile. Odore veramente singolare per una Lepiota, sgradevole, quasi di pesce andato a male. Spore di profilo ellissoidale (ovispore). Osservazione in rosso Congo, a 400×. -
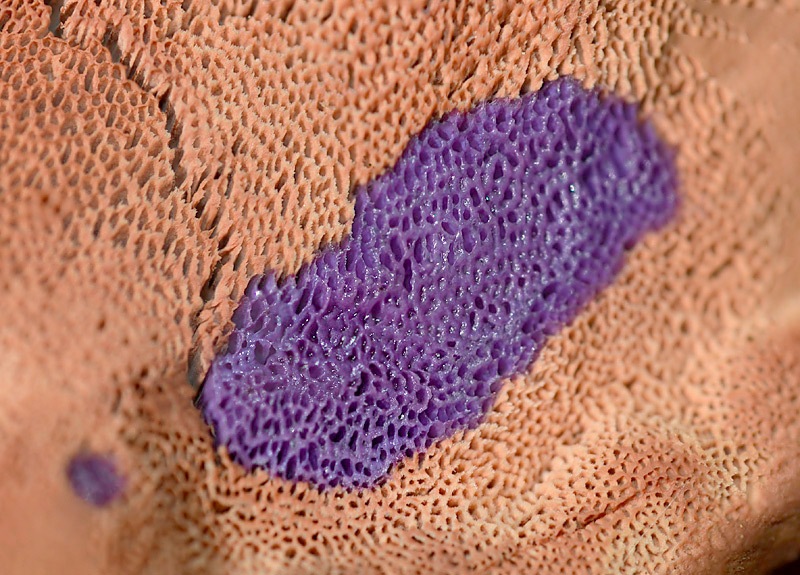
Hapalopilus rutilans (Pers. : Fr.) Murrill 1904
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Velenosi
Hapalopilus rutilans (Pers. : Fr.) Murrill; Regione Toscana; Ottobre 2021; Foto di Tomaso Lezzi. Reazione viola con KOH sia sul cappello (a sinistra) che sui pori (a destra). Particolare della reazione viola sui pori con KOH.