

Alessandro F
-
Numero contenuti
8798 -
Iscritto
-
Ultima visita
Tipo di contenuto
Profili
Forum
Orchidee
Diventa Socio
Calendario
Tutti i contenuti di Alessandro F
-

ODONATA_ 04 - Le ali degli Odonati. Un aiuto per la determinazione
Alessandro F ha risposto alla discussione di Alessandro F in Odonata
4.2.2_ Esempi di indicazioni deducibili dalle osservazioni delle venature supplementari Le venature supplementari, radio supplementare (R-spl) e mediana supplementare (M-spl), ricorrono abbastanza spesso nelle descrizioni di alcune specie di Anisotteri, in quanto fonte di informazioni per individuare il genere di appartenenza; e talvolta utili nel fugare dubbi determinativi nel confronto di specie simili tra loro. In alcuni casi, a seconda dei generi (o delle specie), al di sopra di esse e al di sotto delle venature longitudinali che le precedono, rispettivamente la R-i3 (radiale intercalare) e la mediana, si possono individuare delle celle disposte in file più o meno distinte e ordinate, oppure disposte disordinatamente e numerose o, ancora, presenti ma con diverse celle raddoppiate. Anche l’andamento di R-spl nella sua parte terminale (incurvata verso la punta dell’ala, oppure rivolta più verso il basso) può essere significativo e indirizzare la determinazione verso alcuni generi anziché altri. Così come è interessante osservare se la R-i3 presenta o meno una biforcazione. Solitamente le osservazioni sopra elencate vengono effettuate sull’ala anteriore e dando la preferenza alla R-spl perché talvolta è più evidente della M-spl; ciò non toglie che possano corrispondere anche partendo dalla M-spl e/o anche considerando l’ala posteriore. Qui di seguito alcuni esempi con le relative considerazioni e indicazioni determinative. Per maggior leggibilità, nelle foto che seguono, sono colorate con evidenti tratti lunghi le seguenti venature, a partire dall’alto di ogni ala: Radiale intercalare, R-i3: in giallo (anche nelle sua biforcazione, se esiste); Radiale supplementare, R-spl: in blu; Mediana: in arancione; Mediana supplementare, M-spl: in verde. Nella famiglia Aeshnidae può essere diagnostico conteggiare il numero di file di celle individuabili nelle zone comprese tra R-spl (linea blu) e R-i3 (linea gialla) o, equivalentemente, tra M-spl (linea verde) e mediana (linea arancione). E ciò indifferentemente tra ala anteriore o posteriore, anche se (convenzionalmente) è sufficiente riferire l’osservazione alla sola ala anteriore. Se tale numero varia tra 3 e 5 allora siamo in presenza di una specie appartenente ai generi Aeshna o Anax: solitamente 3-5 file (abbastanza ordinate) per Aeshna e 4-5 file (anche un po' disordinate/confuse) per Anax. Se tale numero risulta uguale a 2 siamo invece in presenza di una specie del genere Boyeria (e quindi di una Boyeria irene, essendo questa l’unica specie di tale genere presente in Italia); se vi si trova una sola fila di celle allora siamo in presenza di una specie del genere Brachytron (e quindi di una Brachytron pratense, essendo questa l’unica specie di tale genere presente in Italia). Nel caso della foto qui sopra, sia osservando l’ala anteriore che quella posteriore, il numero di file (anche se un po’ disordinate) varia da 3 a 4 (le file sono contrassegnate con linee fini e rosse). La scelta può quindi limitarsi a Aeshna o Anax. Ma si vede anche che la R-i3 non presenta biforcazioni e quindi non può essere una Aeshna (che presenta sempre la R-i3 con una biforcazione) ma una Anax (che ha sempre la R-i3 non biforcata). Anche l’andamento terminale di R-spl è indicativo perché, curvando e orientandosi verso la parte distale del pterostigma, denuncia la presenza di una Anax (in Aeshna la R-spl termina la sua corsa dirigendosi assai più in basso). In effetti la foto ritrae un maschio di Anax imperator (Leach, 1815). In questo caso, sia osservando le ali anteriori che quelle posteriori, 3 è il numero di file (assai ben distinguibili e ordinate) riscontrate in quelle particolari zone. La scelta può quindi limitarsi a Aeshna o Anax. Ma si vede anche che la R-i3 si biforca e quindi non può essere una Anax (che ha sempre la R-i3 non biforcata) ma una Aeshna (che presenta sempre la R-i3 con una biforcazione). Osservare anche come la R-spl, nella sua parte terminale, non si incurvi verso l’alto ma sia orientata più verso la zona sottostante alla punta dell’ala. In effetti la foto ritrae un maschio di Aeshna affinis (Vander Linden, 1820). Le due foto precedenti mostrano due maschi, maturo il primo e immaturo il secondo. Si riscontrano in quelle particolari zone al di sopra delle venature supplementari 3 file di celle abbastanza ben ordinate; e inoltre la R-i3 è biforcata e la R-spl continua la sua corsa rivolgendosi in basso. Tutto ciò denuncia una specie del genere Aeshna. Infatti siamo in presenza, in entrambe le foto, di maschi di Aeshna cyanea (Müller, 1764). In questo caso, sia osservando le ali anteriori che quelle posteriori, il numero di file varia tra 3 e 4. La scelta può quindi limitarsi a Aeshna o Anax. Ma si vede anche che le file sono assai ordinate e, soprattutto, che la R-i3 si biforca e che la R-spl, nel suo tratto terminale, non sia rivolta verso l’alto. Quindi non può essere una Anax (che ha sempre la R-i3 non biforcata e la R-spl rivolta verso l’alto nel suo tratto terminale). In effetti la foto ritrae un maschio di Aeshna mixta (Latreille, 1805). 5 file (più o meno disordinate) di celle individuate tra la R-spl e la R-i3, con quest’ultima non biforcata; e con la parte terminale della R-spl che si incurva e si orienta verso la parte distale del pterostigma. Siamo in presenza di una Anax. In effetti la foto ritrae un maschio di Anax ephippiger (Burmeister, 1839). Le osservazioni inerenti la zona tra la R-spl e la R-i3 (ma limitando stavolta l’analisi alla sola ala anteriore) non riguardano soltanto le specie della famiglia Aeshnide ma si estendono anche ad altre famiglie, in particolare alla grande famiglia Libellulidae. Tali osservazioni, integrate poi con altre caratteristiche come: presenza di eventuali macchie colorate alla base delle ali, numero delle celle entro particolari zone (ad esempio in triangolo, sub-triangolo, triangolo anale, campo discoidale, ecc.), forma di alcune zone alari, o altre peculiarità sempre riguardanti le ali, permettono spesso di individuare il genere di appartenenza con un buon margine di sicurezza. Ma di questo ne verrà discusso in altra sede. Qui di seguito riportiamo alcuni esempi che permettono di distinguere due specie di Orthetrum abbastanza simili come Orthetrum coerulescens e Orthetrum brunneum. La colorazione e le caratteristiche morfologiche delle due libellule precedenti potrebbe dare l’impressione di trovarsi di fronte a due esemplari della stessa specie, col dubbio determinativo tra Orthetrum brunneum o Orthetrum coerulescens. Il primo esemplare è un maschio, come si deduce dalla stretta vicinanza delle sue appendici addominali; per l’esattezza è un maschio immaturo visto che a maturità i maschi della famiglia Libellulidae perdono quella colorazione giallognola (tipica invece delle femmine) per assumere tutt’altra colorazione più o meno vistosa. Il secondo esemplare è una femmina (appendici addominali discoste e più o meno parallele tra di loro). Potrebbero quindi rappresentare maschio e femmina della stessa specie. Tuttavia la presenza delle celle “raddoppiate” (segnate con i puntini rossi: 7 celle raddoppiate) tra la R-i3 e la R-spl nella prima foto e, di contro, l’assenza di celle raddoppiate nella seconda foto, ne dichiarano la specie: maschio immaturo di Orthetrum brunneum il primo e femmina di Orthetrum coerulescens la seconda. Statisticamente è stato infatti rilevato che Orthetrum brunneum (in entrambi i sessi) presenta in quella zona un numero di “celle raddoppiate” variabile da 4 a 9 (e raramente, al minimo, 3); mentre Orthetrum coerulescens, sempre statisticamente e in entrambi i sessi, non ne presenta alcuna o, al massimo 2 o 3. Vero è che sono regole empiriche e statistiche e che quindi qualche volta si potrebbe anche incappare nella rara o rarissima eccezione, ma la loro applicazione è ritenuta valida e probante “quasi” sempre, tranne nei casi dubbi (ad esempio trovarsi con 3 celle raddoppiate e non aver la possibilità di osservare altre caratteristiche morfocromatiche). Nel caso di confronto tra maschi maturi i parametri distintivi sono molteplici e riguardano, ad esempio, la fronte (celeste in Orthetrum brunneum, bruna o comunque non celeste in Orthetrum coerulescens) o la parte dorsale del torace (più estesamente e uniformemente celeste in Orthetrum brunneum, raramente così uniformemente colorata in Orthetrum coerulescens ma più spesso brunastra o bruno-cilestrina) o la forma dell’addome (più largo in Orthetrum brunneum, più affusolato in Orthetrum coerulescens) o l'osservazione del pterostigma (più allungato in Orthetrum coerulescens rispetto a quello di Orthetrum brunneum); caratteristiche queste ultime, riguardanti addome e pterostigma, non sempre ben apprezzabili senza avere a disposizione due esemplari in contemporaneo confronto. E, infine, la suddetta “regola delle celle raddoppiate”. Alcuni esempi. Maschio di Orthetrum brunneum; confermato dalla fronte celeste e da 4-5 celle raddoppiate (segnate con i puntini rossi) nella tipica zona tra R-spl e R-i3 Maschio di Orthetrum brunneum; confermato dalla fronte celeste e da 4 (ala sinistra) e 5 (ala destra) celle raddoppiate nella tipica zona tra R-spl e R-i3. Osservare come nelle ali posteriori la "regola delle celle raddoppiate" non fornisca risultato probante, con 3 celle raddoppiate. Maschio di Orthetrum brunneum; confermato dalla fronte celeste e da 7 celle raddoppiate nella tipica zona tra R-spl e R-i3. Maschio di Orthetrum brunneum; confermato dalla fronte celeste e da 5 celle raddoppiate nella tipica zona tra R-spl e R-i3. Osservare come nelle ali posteriori la "regola delle celle raddoppiate" non fornisca risultato coerente; anzi, l'assenza di celle raddoppiate farebbe addirittura pensare a Orthetrum coerulescens. Maschio di Orthetrum brunneum; confermato dalla fronte celeste e da 5 celle raddoppiate nella tipica zona tra R-spl e R-i3. Femmina di Orthetrum coerulescens; ipotesi dovuta all'assenza di celle raddoppiate in quella tipica zona; ipotesi confermata dall'assenza di 2 bande chiare più o meno soffuse sul fianco del torace (presenti invece nelle femmine di Orthetrum brunneum) e dalle brevi tracce scure presenti sul dorso dell'addome (indicate dalle frecce gialle): tracce che nelle femmine di Orthetrum brunneum si presentano solitamente come singoli punti isolati e separati (uno di qua e uno al di là rispetto alla carena dorsale addominale) mentre nelle femmine di Orthetrum coerulescens si presentano solitamente come piccole linee poco o per niente separate tra di loro (in questa foto le si vede collegate da una parte all'altra della carena dorsale addominale). Femmina di Orthetrum coerulescens; ipotesi dovuta all'assenza di celle raddoppiate in quella tipica zona; ipotesi confermata dalle brevi tracce scure presenti sul dorso dell'addome: tracce che nelle femmine di Orthetrum brunneum si presentano solitamente come singoli punti isolati e separati (uno di qua e uno al di là rispetto alla carena dorsale addominale) mentre nelle femmine di Orthetrum coerulescens si presentano solitamente come piccole linee poco o per niente separate tra di loro (in questa foto le si vede collegate da una parte all'altra della carena dorsale addominale). Maschio di Orthetrum coerulescens; ipotesi dovuta all'assenza di celle raddoppiate in quella tipica zona; ipotesi che non può essere confermata dalla fronte che qui non si vede; tuttavia confermata dalle brevi tracce scure presenti sul dorso dell'addome: tracce non sempre visibili nei maschi perché spesso nascoste dallo strato pruinoso celeste, ma se sono visibili si presentano solitamente come singoli punti ben separati in Orthetrum brunneum, mentre nei maschi di Orthetrum coerulescens si presentano solitamente come piccole linee poco o per niente separate tra di loro; anche il pterostigma assai allungato può essere indicativo; così come l'addome ben affusolato. Maschio di Orthetrum coerulescens; stesse considerazioni precedenti, vista la presenza di sole 2 celle raddoppiate in quella zona caratteristica. Altri esempi su come l’osservazione delle celle collocate tra la R-spl (colorata in rosso) e la R-i3 (colorata in verde) permetta di fugare ogni dubbio tra due specie della famiglia Libellulidae, talvolta confuse tra di loro. Crocothemis erythraea (Brullé, 1832) in quella zona presenta due file di celle mentre Trithemis annulata (Palisot de Beauvois, 1807) ne presenta una sola. Sopra: femmina di Trithemis annulata; sotto: femmina di Crocothemis erythraea. Sopra: maschio di Trithemis annulata; sotto: maschio di Crocothemis erythraea. Talvolta può essere utile controllare il numero di file celle presenti al di sotto della R-spl. Come in questo caso in cui il numero di file di celle compreso tra la R-spl e il bordo inferiore dell'ala varia tra 4 e 5. Questa, assieme ad altre caratteristiche, permette di differenziare Sympetrum sanguineum (Müller, 1764) [famiglia Libellulidae] dalla simile Sympetrum depressiusculum che, nella stessa zona, ha molte più celle e disposte di norma in 5-7 file..jpg.9a64ddc303d59299838d355b9c47d982.jpg)
.jpg.6958393e6a9cbfbf31323e27276042cf.jpg)
.jpg.5b8a5fbe86b6d5faffde732e3bacfada.jpg)
.jpg.6d82ecd834f50c0cb1e50721df37bf92.jpg)
.jpg.6ca63da1fa692989ffd71915085e7ae4.jpg)
.jpg.b4fd1eeafc06b9ca684a11af35fe8095.jpg)
.jpg.45128c8e3c72ff687538e961dde8be6b.jpg)
.jpg.dcc9302246df64007105651c054fed24.jpg)
.jpg.2dc2290ba89c40fdd7a789ad0a529726.jpg)
.jpg.581139b36611bef429227b36ca25b1af.jpg)
.jpg.f2431c5f389e659c2fbf3a34e2c3138b.jpg)
.jpg.0d4885f39c754af4510e21e0d35baa54.jpg)
.jpg.df03aba4c4b6adb2103461090c3ffb1b.jpg)
.jpg.7b9361b7d255c9783140fb4c7d141beb.jpg)
.jpg.6fb8df5ca64ab78c87cb0ef1633c620a.jpg)
.jpg.8578464a7afebfc3b1df65fccea2f81c.jpg)
.jpg.5a66350583f923360c1dbed9c3681db6.jpg)
.jpg.97768ba1e17ee600a2921d5fc1ed790d.jpg)
.jpg.4e5c4a52205332f48b14660a9c7a70a4.jpg)
.jpg.2a260f9bcbfeb8fa7514d841919ddf9c.jpg)
.jpg.a59e0f7b9a9900621ba800439ea9dc6e.jpg)

-
ODONATA_ 04 - Le ali degli Odonati. Un aiuto per la determinazione
Alessandro F ha risposto alla discussione di Alessandro F in Odonata
4.2_ Alcune indicazioni diagnostiche deducibili dall'osservazione delle ali degli Anisotteri Più informazioni si posseggono e più è facile circoscrivere la ricerca verso un particolare taxon di appartenenza. L’analisi di alcune venature, come dagli esempi esposti precedentemente, può già fornire alcune buone informazioni. Se a queste ne vengono aggiunte altre riguardanti particolari zone alari (presenza/assenza; loro forma; numero di celle in esse compreso; particolari colorazioni o presenza di macchie colorate) allora la collocazione in un taxon invece che in un altro risulta facilitata. Senza entrare in merito ad una determinazione specie per specie, né pensare di proporre una casistica esaustiva, vediamo qualche esempio su come l’osservazione delle ali degli Anisotteri possa circoscrivere la ricerca. 4.2.1_ Triangoli e loro orientamento. Venature trasverse antenodali delle ali anteriori Tra la base dell’ala, il nodo e il sub-nodo si individuano facilmente sia la venatura costale (o costa) che la venatura radiale (R1; inizialmente unita alla venatura mediana ma che, dopo l’arculo, prosegue da sola). Tra queste due importanti venature longitudinali si trova, più o meno parallela ad esse e posizionata a pari distanza, la venatura subcostale (o subcosta). Rimanendo esclusivamente nell’ambito dell’ala anteriore (ne vedremo poi il motivo), passiamo a considerare sia le brevi venature trasverse (o trasversali) “superiori” che collegano la costa alla subcosta, sia le altrettanto brevi venature trasverse “inferiori” che collegano la subcosta al radio. Di tali venature può tornare utile apprezzare un loro eventuale cambio di spessore, oppure controllare se esiste o meno un buon allineamento tra le “superiori” e le “inferiori”, o anche contare il numero di quelle “superiori”. Di seguito saranno evidenziati, colorati, anche i triangoli presenti sia su ala anteriore che su ala posteriore e sarà indicato con una freccia il loro orientamento. Tutto diventa più chiaro con i seguenti esempi e con le considerazioni che ne derivano. Per brevità scriveremo semplicemente “venature” intendendo, qui, che si tratta sempre di “venature trasverse ante-nodali”. A: sono indicate le venature di maggior spessore e che mostrano un perfetto allineamento tra superiori e inferiori. Tutte le altre venature (come, per esempio, quelle indicate con le frecce in B), sono di spessore minore e inoltre si mostrano più o meno disallineate tra superiori e inferiori; anche il loro numero è diverso essendo le superiori più numerose. In questo caso quello che più conta è la presenza delle venature indicate dalla freccia A; a tutti gli effetti è come considerare non 2 coppie di venature (2 superiori e 2 inferiori) più grosse e ben allineate ma semplicemente soltanto 2 spesse ed evidenti venature che collegano direttamente la venatura costale con quella radiale, e intersecate nella loro metà dalla venatura subcostale. Ai fini della determinazione (rimanendo in un ambito europeo), questa caratteristica delimita la ricerca tra 3 sole famiglie: Aeshnidae, Cordulegastridae e Gomphidae; escludendo automaticamente la possibilità di trovarsi di fronte ad una specie appartenente alle famiglie Libellulidae, Corduliidae o Syntmemistidae. E vale anche il viceversa: l’assenza di tali 2 venature ben allineate, di spessore evidente e superiore alle altre, farà indirizzare la ricerca tra le famiglie Libellulidae, Corduliidae o Syntmemistidae. Le stesse considerazioni valgono anche per quanto riguarda l’orientamento dei triangoli: se il loro angolo acuto “guarda”, sia nell'ala anteriore che in quella posteriore, verso la punta dell’ala stessa (come nel caso di questa foto), allora si può delimitare la ricerca tra le famiglie Aeshnidae, Cordulegastridae e Gomphidae, escludendo automaticamente la possibilità di trovarsi di fronte ad una specie appartenente alle altre 3 famiglie. E vale il viceversa quando si riscontra che l’orientamento dei triangoli differisce passando dall’ala anteriore a quella posteriore. La foto ritrae in effetti un esemplare della famiglia Aeshnidae, un maschio di Aeshna cyanea (Müller, 1764). A: sono indicate le 2 coppie di venature di maggior spessore e che mostrano un perfetto allineamento tra superiori e inferiori. Tutte le altre venature (come, per esempio, B), sono di spessore minore e inoltre si mostrano più o meno disallineate tra superiori e inferiori; anche il loro numero è diverso essendo le superiori più numerose. I triangoli mostrano evidentemente lo stesso orientamento. Ne segue quanto osservato più sopra. La foto ritrae in effetti un esemplare della famiglia Aeshnidae, un maschio di Aeshna affinis (Vander Linden, 1820). A: sono indicate le 2 coppie di venature di maggior spessore e che mostrano un perfetto allineamento tra superiori e inferiori. Tutte le altre venature sono di spessore minore e inoltre si mostrano più o meno disallineate tra superiori e inferiori; anche il loro numero è diverso essendo le superiori più numerose. I triangoli hanno lo stesso orientamento. Ne segue quanto osservato più sopra. La foto ritrae in effetti un esemplare della famiglia Aeshnidae, un maschio di Aeshna mixta (Latreille, 1805). A: sono indicate le 2 coppie di venature di maggior spessore e che mostrano un perfetto allineamento tra superiori e inferiori. Tutte le altre venature sono di spessore minore e inoltre si mostrano più o meno disallineate tra superiori e inferiori; anche il loro numero è diverso essendo le superiori più numerose. I triangoli hanno lo stesso orientamento. Ne segue quanto osservato più sopra. La foto ritrae in effetti un esemplare della famiglia Cordulegastridae, un maschio di Cordulegaster boltonii (Donovan, 1807). A: sono indicate le 2 coppie di venature di maggior spessore e che mostrano un perfetto allineamento tra superiori e inferiori. Tutte le altre venature sono di spessore minore e inoltre si mostrano più o meno disallineate tra superiori e inferiori; anche il loro numero è diverso essendo le superiori più numerose. I triangoli hanno lo stesso orientamento. Ne segue quanto osservato più sopra. La foto ritrae in effetti un esemplare della famiglia Gomphidae, un maschio di Onychogomphus forcipatus subsp. unguiculatus (Linnaeus, 1758). A: sono indicate le 2 coppie di venature di maggior spessore e che mostrano un perfetto allineamento tra superiori e inferiori. Tutte le altre venature sono di spessore minore e inoltre si mostrano più o meno disallineate tra superiori e inferiori; anche il loro numero è diverso essendo le superiori più numerose. I triangoli hanno lo stesso orientamento. Ne segue quanto osservato più sopra. La foto ritrae in effetti un esemplare della famiglia Aeshnidae, un maschio di Anax imperator (Leach, 1815). Le frecce rosse indicano le 2 coppie di venature di spessore nettamente superiore a tutte le altre; pur non potendo osservare al meglio se esiste allineamento o meno tra le superiori e le inferiori, questo basta per indicare un esemplare appartenente a una delle famiglie Aeshnidae, Cordulegastridae o Gomphidae. Anche i triangoli sono orientati nello stesso modo, il che conferma l’appartenenza ad una delle suddette famiglie. La foto ritrae in effetti un esemplare della famiglia Aeshnidae, un maschio di Anax ephippiger (Burmeister, 1839). Stavolta, tra la base dell’ala anteriore e il nodo, le venature trasverse (sia superiori che inferiori) non mostrano alcun aumento di spessore ma sono tutte simili. Inoltre sono abbastanza allineate tra superiori e inferiori in modo da potersi considerare come “uniche venature trasverse” che collegano direttamente la venatura costale con quella radiale, e intersecate nel mezzo dalla venatura subcostale. In questo caso tali venature “idealmente unificate tra superiori e inferiori” si diranno convenzionalmente “complete” e saranno, appunto, trattate come "uniche". Ciò è in definitiva sufficiente per escludere l’appartenenza a una delle famiglie Aeshnidae, Cordulegastridae o Gomphidae; viceversa la scelta dovrà cadere tra le famiglie Libellulidae, Corduliidae o Syntmemistidae. La stessa identica considerazione si ottiene osservando l’orientamento dei triangoli: nell’ala anteriore il triangolo mostra l’angolo acuto orientato verso la parte inferiore dell’ala, mentre nell’ala posteriore l’angolo acuto del triangolo è diversamente orientato e guarda verso la punta dell’ala stessa; ciò non può che indirizzare la ricerca verso le famiglie Libellulidae, Corduliidae o Syntmemistidae. In questi casi può essere utile anche contare il numero delle venature trasverse (senza più pensarle suddivise tra superiori e inferiori) perché tale numero può dare indicazioni (di massima, ma non assolute) anche sul genere di appartenenza. La foto ritrae in effetti un esemplare della famiglia Libellulidae, un maschio di Libellula depressa (Linnaeus, 1758); il conto delle sue venature trasverse antenodali “complete” deducibili dall’osservazione della foto è di 15 per l’ala destra e 16 per l’ala sinistra. Ciò non è né un’eccezione né una malformazione riguardante le ali. Non sempre, anche considerando la complessità delle loro strutture, le ali anteriori mostrano una perfetta e assoluta simmetria l’una con l’altra, così come accade spesso tra le ali posteriori. Stesse osservazioni e conclusioni fatte nel caso precedente. La foto ritrae in effetti un esemplare della famiglia Corduliidae, una femmina di Somatochlora meridionalis (Nielsen, 1935); il conto delle sue venature trasverse antenodali “complete”, deducibili dall’osservazione della foto della sola ala destra, è di 8. Foto di Massimo Squarcini. Stesse osservazioni e conclusioni fatte nei 2 casi precedenti. La foto ritrae in effetti un esemplare della famiglia Libellulidae, un maschio di Orthetrum cancellatum (Linnaeus, 1758); il conto delle sue venature trasverse antenodali “complete” deducibili dall’osservazione della foto è di 14 per l’ala destra e di 15 per l’ala sinistra. Anche stavolta l’orientamento dei triangoli cambia passando dall’ala anteriore a quella posteriore. Ciò esclude l’appartenenza a una delle famiglie Aeshnidae, Cordulegastridae e Gomphidae; mentre indirizza l’indagine tra le famiglie Libellulidae, Corduliidae o Syntmemistidae. Tuttavia si osserva che le venature trasverse sono di pari spessore e anche ben allineate tra superiori e inferiori (frecce gialle), tutte tranne (!) quelle più vicine al nodo che sono soltanto superiori (freccia blu) e non oltrepassano la venatura subcostale; tali venature si dicono, convenzionalmente, “incomplete” per caratterizzarle da quelle dette “complete” viste nei 3 esempi precedenti. In questi casi (ricordiamo che stiamo sempre parlando di ambito europeo) tale osservazione circoscrive ancora di più la ricerca che, escludendo le famiglie Corduliidae o Syntmemistidae le cui specie hanno venature trasverse antenodali tutte “complete”, viene indirizzata verso la famiglia Libellulidae. Ma in tale famiglia i generi Orthetrum e Libellula annoverano solo specie con venature trasverse antenodali “complete” (tranne qualche sporadica eccezione) e quindi circoscrivendo ancor più la ricerca tra i generi Sympetrum, Trithemis, Crocothemis (i più frequenti da incontrare) o Brachythemis, Pantala, Zygonyx (assai più rari da incontrare in Italia, e relegati essenzialmente alle isole maggiori, Sicilia e Sardegna). In questi casi il conto delle venature trasverse antenodali viene convenzionalmente fatto contando il numero delle “venature complete” più quella “incompleta” a cui si attribuisce il "valore di ½”. La foto ritrae in effetti un maschio di Sympetrum striolatum (Charpentier, 1840); il conto delle sue venature trasverse antenodali deducibili dall’osservazione della foto è di 6 e ½ per entrambe le ali. Anche stavolta l’orientamento dei triangoli cambia passando dall’ala anteriore a quella posteriore. Ciò esclude l’appartenenza a una delle famiglie Aeshnidae, Cordulegastridae e Gomphidae; mentre indirizza l’indagine tra le famiglie Libellulidae, Corduliidae o Syntmemistidae. Tuttavia si osserva che le venature trasverse sono di pari spessore e ben allineate tra superiori e inferiori (frecce viola), tutte tranne (!) quelle più vicine al nodo che sono soltanto superiori (frecce blu) e non oltrepassano la venatura subcostale; si tratta quindi, come visto nel caso precedente, delle venature dette convenzionalmente “incomplete”. Si osserva anche che le venature antenodali delle ali posteriori (quantomeno per l’ala sinistra) sono tutte “complete” (freccia nera indicante l’ultima venatura antenodale, completa). La foto ritrae in effetti ancora un maschio di Sympetrum striolatum (Charpentier, 1840); il conto delle sue venature trasverse antenodali è di 6 e ½ per entrambe le ali. Stesse considerazioni del caso precedente. Osservare che nelle ali posteriori le venature trasverse antenodali siano tutte “complete”, comprese anche quelle (frecce gialle) più vicine al nodo. Mentre è evidente, osservando le sole ali anteriori, come siano presenti le venature “incomplete” (frecce blu) in prossimità del nodo. La foto ritrae in effetti un maschio di Trithemis annulata (Palisot de Beauvois, 1807); il conto delle sue venature trasverse antenodali deducibili dall’osservazione della foto è di 9 e ½ per l’ala sinistra e 10 e ½ per quella destra. Stesse considerazioni del caso precedente. Osservare che nell'ala posteriore le venature trasverse antenodali (frecce verdi) sono tutte “complete”. Mentre è evidente, osservando l'ala anteriore, come sia presenta la venatura “incompleta” (freccia blu) in prossimità del nodo. La foto ritrae in effetti un maschio immaturo di Sympetrum striolatum (Charpentier, 1840); il conto delle sue venature trasverse antenodali deducibili dall’osservazione della foto è di 6 e ½. Stesse considerazioni del caso precedente. Osservare che nelle ali posteriori le venature trasverse antenodali siano tutte “complete”. Mentre è evidente, osservando le sole ali anteriori, come siano presenti le venature “incomplete” (frecce blu) in prossimità del nodo. La foto ritrae in effetti un maschio immaturo di Crocothemis erythraea (Brullé, 1832); il conto delle sue venature trasverse antenodali deducibili dall’osservazione della foto è di 9 e ½ per ciascuna ala anteriore. Stesse considerazioni del caso precedente. Osservare che nelle ali posteriori le venature trasverse antenodali sono tutte “complete”. Mentre, osservando le sole ali anteriori, è evidente la presenza delle venature “incomplete” (frecce blu) in prossimità del nodo. La foto ritrae in effetti un maschio di Crocothemis erythraea (Brullé, 1832); il conto delle sue venature trasverse antenodali deducibili dall’osservazione della foto è di 9 e ½ per ciascuna ala anteriore. Stesse considerazioni del caso precedente. Osservare che nelle ali posteriori le venature trasverse antenodali sono tutte “complete”, 5 per ogni ala. Mentre, osservando le sole ali anteriori, è evidente la presenza delle venature “incomplete” (frecce blu) in prossimità del nodo. La foto ritrae in effetti un maschio di Sympetrum meridionale (Sélys, 1841); il conto delle sue venature trasverse antenodali deducibili dall’osservazione della foto è di 6 e ½ per ciascuna ala anteriore. Dagli ultimi esempi riportati è evidente che l’osservazione sulla presenza o meno delle venature “incomplete” va compiuta esclusivamente sulle ali anteriori perché le ali posteriori non darebbero indicazioni corrette. Inoltre può accadere, e non sono casi rarissimi, di incontrare esemplari in cui l’osservazione delle venature trasverse antenodali delle ali anteriori sembra contraddire le regole suddette o comunque lasciare qualche dubbio. D’altra parte qui si tratta di venature veramente piccole e l’eccezione può benissimo essere giustificata da un incidente che ha fatto sparire una venatura, o da un problema durante il passaggio da ninfa a imago o semplicemente da una variante soggettiva. Viceversa l’orientamento del triangolo (struttura ben più complessa e particolare rispetto a una piccola venatura) è assolutamente indicativo, e quindi di più probante aiuto diagnostico ai fini determinativi, tranne evidenti malformazioni delle ali. Vediamo qualche eccezione a proposito delle venature antenodali (eccezione che comunque e in definitiva... conferma la regola). Femmina di Crocothemis erythraea (Brullé, 1832). I triangoli sono orientati come ci si aspetta da un Libellulidae o da un Corduliidae o Syntmemistidae; ma questi ultimi 2 sono però da escludere perché dovrebbero avere colorazioni corporee nettamente diverse (più metalliche e più sul verdognolo). Inoltre e partendo prima di tutto dalla visione delle appendici addominali che denotano una femmina, si osserva quanto segue: forma dell’addome, colorazione del corpo associata a una evidente stria chiara sul dorso del torace tra le basi delle ali, colorazione tipica delle “macchie” alla base delle ali, tipica colorazione degli occhi… osservando tutto ciò, non ci sono dubbi interpretativi: è una femmina di Crocothemis erythraea. Tuttavia se analizziamo le venature trasverse antenodali se ne contano 9, e tutte complete, sull’ala di destra mentre in quella di sinistra ve ne sono 8 complete più una “indecisa” in quanto non ben allineata e neanche "scolasticamente" incompleta. Evidentemente si tratta di una eccezione che conferma la regola; eccezione nata così, senza incidenti di sorta, considerando che l’esemplare è molto giovane a giudicare dalla ali ancora perfettamente integre e senza la minima ammaccatura o lacerazione. Maschio di Orthetrum coerulescens (Fabricius, 1798). I triangoli sono orientati come ci si aspetta da un Libellulidae o da un Corduliidae o Syntmemistidae; ma questi ultimi 2 sono però da escludere perché dovrebbero avere colorazioni corporee nettamente diverse (più metalliche e più sul verdognolo). Partendo poi dalla colorazione dell’addome (dovuta alla presenza di una diffusa pruinosità azzurrognola) e dalla posizione delle appendici addominali si deduce che è un maschio di Orthetrum; altri maschi simili e congeneri possono essere Orthetrum cancellatum (da escludere perché ha addome più grande e meno cilindrico, pterostigma neri, così come sono nerastri gli ultimi segmenti addominali), Orthetrum albistylum (da escludere perché ha appendici addominali bianche, oltre che pterostigma neri, così come sono nerastri gli ultimi segmenti addominali), e Orthetrum brunneum. Con quest’ultimo, in effetti, il dubbio determinativo può essere in agguato. Ma Orthetrum brunneum ha colorazione più diffusamente azzurrognola anche sul dorso del torace, ha pterostigma più corti, addome più largo e (ecco che l’analisi delle ali torna molto utile!) presenta solitamente da 4 a 9 celle raddoppiate nella zona alare (vedere la zona colorata indicata dalle frecce gialle) compresa tra la R-i 3 (venatura intercalare, posta al di sotto della R 3) e la R-spl (venatura radiale supplementare), mentre Orthetrum coerulescens non ne presenta alcuna o, al massimo e raramente, ne può presentare fino a 2 o 3. Dalla nostra foto, in ciascuna di quelle particolari zone, vediamo solo 2 celle raddoppiate. Quindi l’esemplare in foto è indubbiamente un maschio di Orthetrum coerulescens. E se osserviamo le venature antenodali delle ali anteriori osserviamo che le venature più vicine ai nodi sono decisamente complete, ma vediamo anche delle piccole incongruenze: con A sono segnalate due venature superiori ma una sola inferiore; con B si indicano venature superiori per niente allineate con le inferiori. Tuttavia anche queste piccole deviazioni dalla regola possono considerarsi eccezioni che talvolta si presentano. Evidente maschio di Orthetrum albistylum (Sélys, 1848); le appendici addominali ravvicinate e bianche non lasciano dubbi. E quella venatura “superiore” mancante (indicata dalla freccia celeste) non può che essere una… “svista” del DNA (oppure una venatura perduta in seguito a qualche accidente). E non si può certo pensare di inserire l’esemplare della foto in qualche altra famiglia. Osservare anche come i triangoli abbiano orientamento che cambia passando dalle ali anteriori a quelle posteriori. Maschio di Libellula quadrimaculata (Linnaeus, 1758). Forse la libellula più facile da determinare grazie alle macchie scure presenti nei pressi del nodo di ciascuna ala (come appropriatamente dichiara il suo nome specifico). Appartenendo al genere Libellula, dovrebbe “scolasticamente” avere le venature trasverse antenodali tutte abbastanza allineate e complete; ma in A c’è solo la venatura superiore, in B l’allineamento non è perfetto e, soprattutto, in C la venatura è interrotta a metà, cioè è incompleta. Tuttavia la determinazione è ineccepibile; e anche questa eccezione… conferma la regola. D’altra parte se analizziamo i triangoli osserviamo che loro le regole le hanno sempre rispettate. Maschio di Libellula depressa (Linnaeus, 1758) [famiglia Libellulidae]; a destra con tutte le venature complete; a sinistra con l'ultima venatura incompleta (freccia). Il che non comporta alcun problema sulla determinazione..jpg.138cd65e829c8555c3774975d378cbee.jpg)
.jpg.cfe4e78dc865644c631770d7039516f3.jpg)
.jpg.ad6963df40eec24a60ac1ffb89200c9a.jpg)
.jpg.42427bbdeabd749b59d93f381f381a09.jpg)
.jpg.5c3f6f4d661c4c06557db037aae1d784.jpg)
.jpg.375f2da4834ec51cd1e9c69114c30f99.jpg)
.jpg.3f158266da9b91cc1b04150856133596.jpg)
.jpg.212299eb0430eb92efd9ffcfeab38864.jpg)
.jpg.b0a66977b46977bf791a72d7fd75941e.jpg)

.jpg.5b5c64ef225c200df9c61802ac17406c.jpg)
.thumb.jpg.45fbe53475160ade09318908785ef295.jpg)
.jpg.d192f871ec06e72a4fc60990cfe2e028.jpg)
.jpg.b66c7085c2e5481b7019917cdb97d7a1.jpg)
.jpg.ac7bac2fbd4a8666d8e02aed031d748e.jpg)
.jpg.bfebda759641b5df74e5afaeb163cd04.jpg)
.jpg.c076119c7f70c75588ea680d4a9b41b8.jpg)
.jpg.c1c5a6919c14df702d7c14fab1a36c58.jpg)
.jpg.f979cd2d567654d4f01693b522895204.jpg)
.jpg.399b5587e756d61a5e55d755f1039b57.jpg)
.jpg.98b791d0ea825559474d939673f3bf7a.jpg)

.jpg.d60ae8efcf95a87e426209c853d976a1.jpg)
-
ODONATA_ 04 - Le ali degli Odonati. Un aiuto per la determinazione
Alessandro F ha postato un topic nell'area Odonata
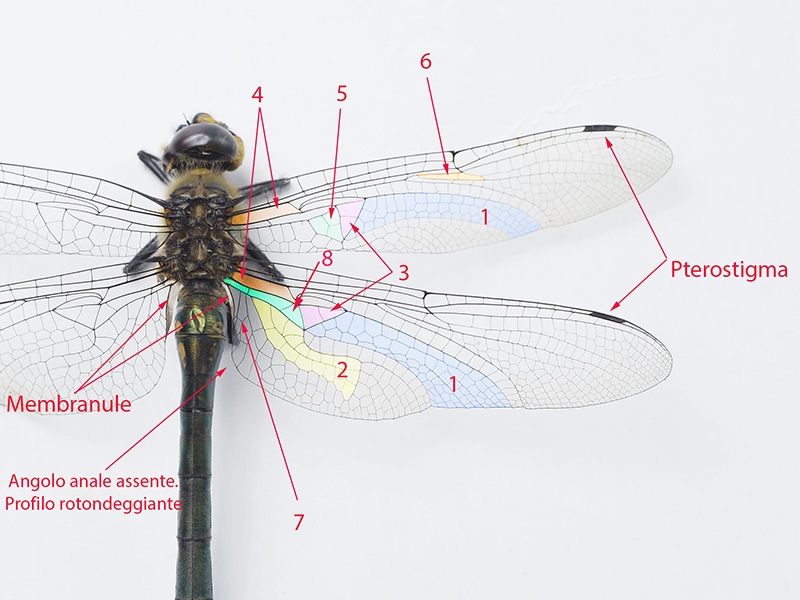
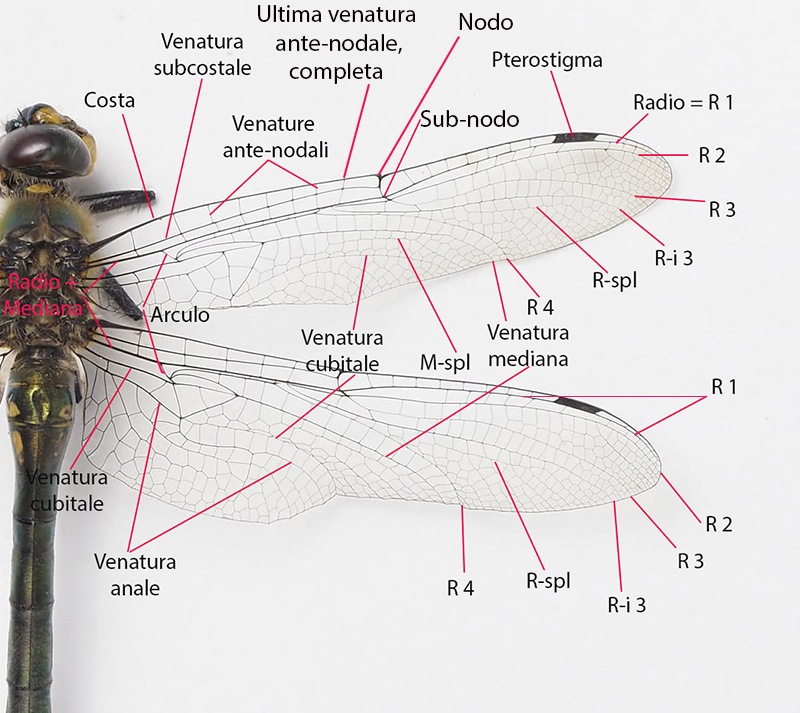
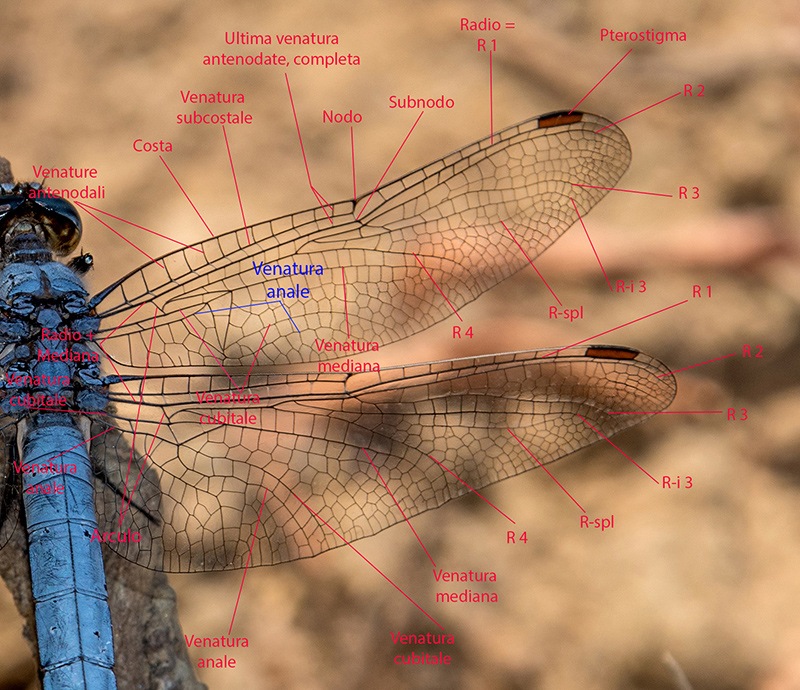
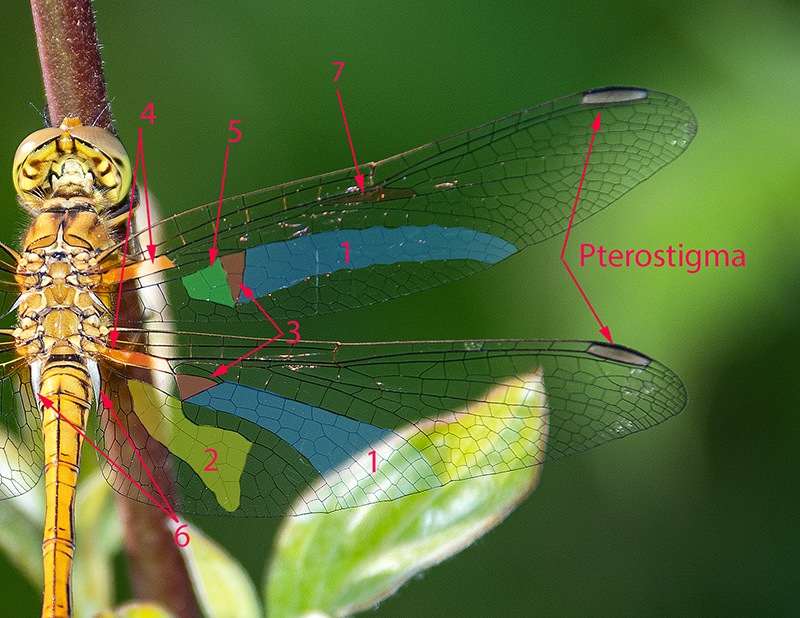
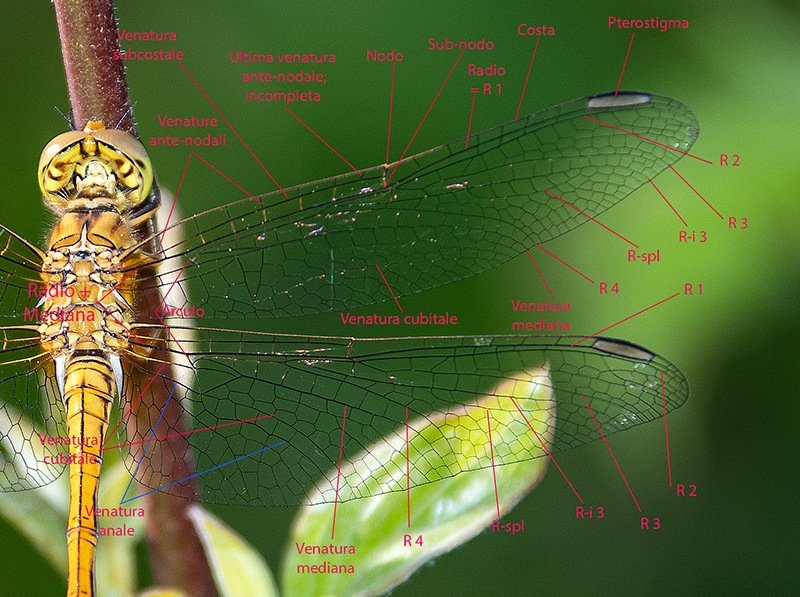
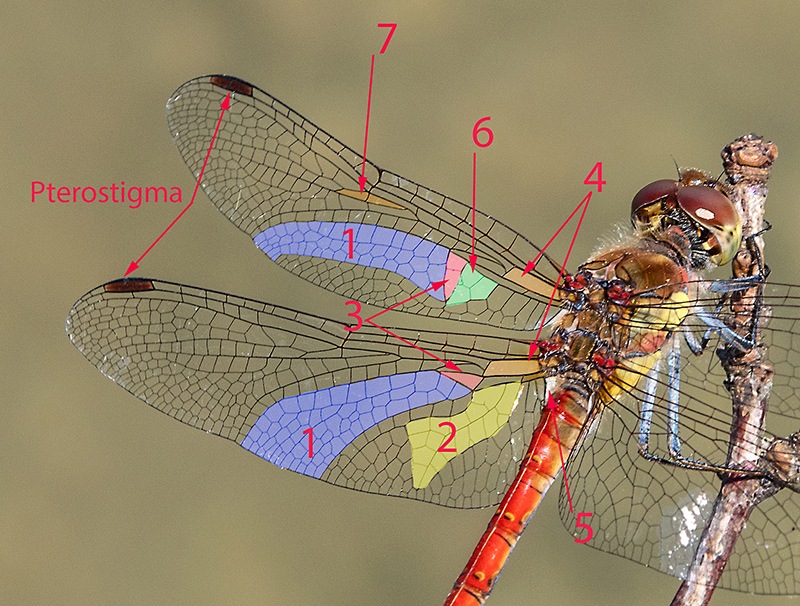
Determinare genere e specie degli Odonati presenti in Italia non richiede solitamente né una lunga esperienza né un'attrezzatura sofisticata come accade, per esempio, nell'indagine di alcune specie di funghi (talvolta con necessità di microscopia o, addirittura, analisi del DNA) o per certe farfalle e falene (analisi accurata dei genitali). Forse ciò deriva dal fatto che il numero di specie di libellule attualmente reperibili in Italia non supera le 100 unità; forse dipende dalla loro colorazione e dai loro disegni spesso così indicativi o dalla territorialità talvolta esclusiva di determinate regioni o habitat; sicuramente è favorita dalla possibilità di fotografarle con una attrezzatura sufficientemente adeguata e di controllare poi con tutta calma le loro caratteristiche strutturali e cromatiche. Tra queste caratteristiche non può mancare lo studio di uno dei loro organi più appariscenti: le ali. A una prima e superficiale occhiata, le ali delle libellule sembrano tutte molto simili tra di loro. Vedremo invece come tale idea di uniformità sia errata e come, viceversa, l'analisi delle ali rappresenti un aiuto, meglio se supportato da altre osservazioni, per distinguere con relativa facilità le famiglie di appartenenza, talvolta anche i generi o il sesso. Ciò vale in primo luogo per le libellule più grandi e appariscenti (sottordine Anisoptera) sia per la relativa grandezza delle loro ali sia, soprattutto, per la maggior facilità di osservazione rispetto alle ali delle specie del sottordine Zygoptera. Infatti gli Anisotteri, quando sono posati, tengono generalmente le 4 ali ben aperte e perpendicolari rispetto all'asse testa-torace-addome (come le pagine aperte di un libro), mentre gli Zigotteri, esclusi quelli della famiglia Lestidae, da posati tengono le 4 ali riunite assieme, in senso longitudinale sopra all'addome e quindi difficili (se non impossibili) da osservare separatamente una dall'altra. Sempre a proposito degli Anisotteri e disponendo di opportune immagini è possibile constatare che le ali non sono conformate "a superficie piatta", come sembrerebbe a prima vista; viceversa presentano, grazie ad alcune venature portanti, zone più o meno rialzate; venendo così ad essere conformate con una sorta di lieve tridimensionalità, soprattutto in vicinanza delle base delle ali. Caratteristica che sicuramente favorisce la loro aerodinamicità. Femmina di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]; Inquadrando l'ala anteriore di infilata e da sotto, è possibile rendersi conto di come le ali, soprattutto degli Anisotteri, non siano affatto piatte ma, viceversa, strutturate tridimensionalmente; con le venature più grandi e robuste che creano delle vere e proprie zone rialzate/abbassate. Evidentemente tale conformazione porta a degli innegabili vantaggi aerodinamici. Orthetrum cancellatum (Linnaeus, 1758) [sottordine Anisoptera; famiglia Libellulidae]. Anche da questa inquadratura si può notare come le ali non siano del tutto piatte ma, almeno verso la zona "costale", presentino un susseguirsi di zone rialzate e abbassate. Una sorta di lieve tridimensionalità che sicuramente favorisce e accentua la loro perfetta aerodinamica. 4.1_ Analisi e studio delle ali degli Anisotteri Per condurre un'analisi appropriata delle ali occorre prendere confidenza con le loro venature (o nervature) e con alcune delle particolari zone di raggruppamento di celle che si vengono a formare; sarà necessario anche appropriarsi della corrispondente nomenclatura. Analizzando un "modello campione" come la grande Anax imperator si possono notare 5 principali venature longitudinali di discreto spessore che emergono dal torace, direttamente innestate nei muscoli di mesotorace (per le ali anteriori) e di metatorace (per le ali posteriori). Si tratta, partendo dalla zona superiore, della grande "venatura costale" (o, brevemente, "costa"), delle due venature "radiale" (detta brevemente "radio") e "mediana" e che inizialmente sono a stretto contatto l'una con l'altra separandosi successivamente a livello del cosiddetto "arculo"; segue poi una "venatura cubitale" e infine una "venatura anale". Chiude il tutto una più esile venatura costale inferiore che non è tuttavia innestata nella muscolatura ma sorge direttamente dalla zona basale della venatura anale, dopo una cella opaca e non trasparente detta "membranula". Maschio di Anax imperator (Leach, 1815) [famiglia Aeshnidae]. Ciò vale per tutte le ali sia anteriori che posteriori. Anticipiamo però, come vedremo meglio in seguito, che tra le ali anteriori e quelle posteriori vi sono differenze morfologiche e strutturali abbastanza evidenti. Le 5 venature suddette si prolungano poi longitudinalmente oppure cambiano orientamento nel loro percorso lungo le ali, qualcuna anche biforcandosi. Ci sono altre venature interessanti da osservare, come la "venatura subcostale" collocata tra la costa e il radio, o come le due "venature supplementari" situate più o meno al centro delle ali ma non collegate direttamente alle precedenti venature né a loro eventuali biforcazioni. Tutte le venature suddette, intersecate da molte altre decisamente più corte e di esile spessore, sia longitudinali che trasversali, contribuiscono a formare una fitta rete di piccole zone (dette "celle" o "cellule") membranose e molto sottili, trasparenti e più o meno colorate a seconda della specie. Le venature rappresentano perciò l'impalcatura generale delle ali ma sono da considerare anche come veri e propri condotti tubuliformi entro cui scorre l'emolinfa (quanto meno all'inizio della vita da adulto, in modo da facilitare la distensione dell'ala in tutta la sua ampiezza) e in cui possono trovarsi sia i nervi che alcune trachee respiratorie. Iniziamo ad osservare più da vicino l'ala di un Anisottero, analizzandone venature e zone caratteristiche a partire dalla costa e scendendo verso la parte inferiore. Vedremo qui di seguito in che modo queste venature si differenziano e come contribuiscono a delimitare alcune particolari regioni spesso di un certo aiuto per l'identificazione delle famiglie o delle specie. Prenderemo confidenza anche con i loro nomi e con i simboli che le contraddistinguono. Lungo la costa, più o meno a metà strada tra la base e la punta dell'ala, si nota un punto in cui avviene una sorta di cambio di direzione, corrispondente anche a una venatura trasversale breve ma di sezione apprezzabile. Tale punto è detto "nodo". Le brevi venature trasversali osservabili subito al di sotto della costa e collocate tra il nodo e la punta dell'ala sono dette "venature post-nodali"; molto più interessanti da osservare sono però le "venature ante-nodali", collocate cioè tra il nodo e la base dell'ala, perché dal loro numero e dal confronto/corrispondenza con le altre brevi venature trasversali poste al di sotto della venatura subcostale si ricavano alcune indicazioni interessanti ai fini determinativi. Poco al di sotto del nodo, in una zona dell'ala in cui alcune venature radiali si biforcano vi è il "sub-nodo". Proseguendo la costa verso la punta dell'ala si osserva una grande cella, ispessita, allungata e colorata diversamente dalle altre, detta pterostigma. Tale cella ha funzioni aereodinamiche costituendo una sorta di rinforzo strutturale in una zona in cui l'ala sottostà ad un notevole sforzo, impedendo perciò che la punta dell'ala si ripieghi deformandosi in maniera anomala durante il volo. Al di sotto della costa e parallelamente ad essa si nota la "venatura subcostale" (o subcosta) la cui corsa termina al di sotto del nodo. Come già detto in precedenza, alla base dell'ala e sotto alla subcosta si notano le due venature radio e mediana che si separano poco dopo in corrispondenza dell'arculo. La venatura radiale è detta così perché con le sue biforcazioni si irradia su gran parte della superficie alare; biforcazioni simboleggiate con R1, R2, R3, R4; con R1 viene indicata la venatura del radio che dopo l'arculo "va a diritto", oltrepassa la zona del nodo, costituisce la delimitazione inferiore dello pterostigma e termina sul bordo della punta dell'ala; R2, R3 e R4 sorgono assieme dall'arculo per biforcarsi e separarsi successivamente (R2 e R3 si separano all'altezza del sub-nodo), con R2 che termina verso la punta dell'ala, R3 verso la punta ma in zona inferiore, mentre R4 termina verso la metà della parte inferiore dell'ala. A queste si aggiunge una "venatura intercalare" o "venatura interposta" (indicata con R-i3 o con iR3, oppure con altra simbologia appropriata e comunque atta a ricordare che tale venatura si colloca vicino alla R3) che segue quasi parallelamente la R3 rimanendovi al di sotto. La R-i3, a seconda del genere di appartenenza, può o meno biforcarsi dando così altre indicazioni diagnostiche. Si nota poi una venatura particolare che non ha un inizio ben definito e che si colloca nella zona alare tra la R-i3 e la R4; tale venatura è detta "radio supplementare" ed è indicata brevemente con R-spl. Anch'essa è spesso rammentata nella descrizione di alcune famiglie o generi di Anisotteri. Continuando l'analisi dell'ala e scendendo verso la zona inferiore incontriamo di nuovo la "venatura mediana" che, scendendo dall'arculo e cambiando orientamento, va a terminare sul bordo inferiore dell'ala nei pressi della R4. A seguire e spostandoci ancor più verso la base inferiore dell'ala troviamo i proseguimenti della "venatura cubitale" e della "venatura anale", con quest'ultima talvolta non particolarmente marcata nell'ala anteriore ma sempre evidente nell'ala posteriore. Nella zona al di sotto della mediana e al di sopra della cubitale si nota un'altra venatura supplementare, detta "venatura mediana supplementare" e indicata con M-spl, con un andamento più o meno incerto. Quanto descritto fin qui per le venature e lo pterostigma vale per tutte le 4 ali, sia anteriori che posteriori. Tuttavia riprendendo la visione dell'ala posteriore si può notare che essa è assai più grande dell'ala anteriore soprattutto in larghezza, contenendo quindi un numero di celle nettamente superiore. Ciò è ancor più evidente osservando la parte inferiore dell’ala posteriore in cui l’ala stessa si allarga soprattutto in vicinanza dell’addome; in questa zona può o meno manifestarsi il cosiddetto “angolo anale” ove il bordo dell’ala subisce un brusco cambio di direzione. L’angolo anale è caratteristico esclusivamente dei maschi di alcune specie, mentre è sempre assente in tutte le femmine. In sua assenza l'ala si mostra arrotondata in quella zona. Passiamo ora ad alcune "regioni" alari particolari e spesso significative ai fini diagnostici, individuabili grazie ad alcune venature più spesse o marcate che ne costituiscono il perimetro. Tra le regioni più evidenti, in base alla specie, vi sono: "triangolo", "campo discoidale", "campo anale", "spazio mediano", "triangolo anale" (caratteristico dei maschi di alcune specie e mai delle femmine), "membranula", "sub-triangolo", "pterostigma" (di cui si è già detto), "campo sub-nodale", "spazio cubitale", ecc. La loro collocazione sarà chiarita (più che con la descrizione a parole) osservando alcune delle seguenti foto in cui, alla visione oggettiva delle ali segue la visione con le indicazioni per le venature e la successiva visione delle aree significative. Osservando e confrontando a varie riprese le ali "al naturale" con le altre corredate dalle indicazioni strutturali e nomenclaturali si dovrebbe in poco tempo imparare sia a riconoscere le varie strutture che a prendere confidenza con la relativa terminologia. Maschio di Aeshna mixta (Latreille, 1805) [famiglia Aeshnidae]. Venature caratteristiche dell'ala anteriore; e pterostigma. Venature caratteristiche dell'ala posteriore; e pterostigma. Alcune regioni caratteristiche: 1. Campo discoidale (compreso tra il lato inferiore del triangolo, il bordo inferiore dell'ala, la venatura R4 e la venatura cubitale; presente in tutte le ali). 2. Campo anale (si tratta di una netta zona presente solo nelle ali posteriori, rispetto al triangolo si trova spostata più verso la base). 3. Triangolo (una piccola ma evidente zona di forma triangolare, contenente una o più celle, e collocata vicino alla base di ciascuna delle 4 ali). Osservare come in entrambe le ali di questa specie (anteriore e posteriore) i triangoli presentino l'angolo acuto orientato verso la punta dell'ala. 4. Spazio mediano (si trova tra la base dell'ala e l'arculo; privo di venature interne, e quindi costituito da una singola cella, tranne in qualche specie). 5. Membranula (una membrana non trasparente come le altre celle, collocata alla base dell'ala tra la venatura anale e la costa inferiore; si nota anche come la membranula sia più grande ed evidente nelle ali posteriori). 6. Triangolo anale (presente solo nei maschi di alcune specie; collocato, ove esiste, a contatto con la membranula). 7. Sub-triangolo (regione dal profilo più o meno triangolare, contenente una o più celle; collocata di fianco al triangolo, dalla parte che guarda la base dell'ala; solitamente è più indicativo ai fini determinativi quello dell'ala anteriore). 8. Campo sub-nodale (piccola zona a forma di triangolo allungato, collocata al di sotto del sub-nodo; contenente una o più celle). 9. Angolo anale (presente solo nei maschi di alcune specie). 10. Spazio cubitale, collocato in entrambe le ali tra la loro base e il triangolo. Spesso, nelle descrizioni delle varie specie viene preso in considerazione soltanto quello dell'ala posteriore. L'osservazione delle foto precedenti (e successive) mostra anche come le due ali siano sostanzialmente diverse sia in grandezza (più ampia e larga l'ala posteriore) sia per alcune zone particolari, come ad esempio il "campo anale", presente esclusivamente nelle ali posteriori. Maschio di Anax imperator (Leach, 1815) [famiglia Aeshnidae]. Venature caratteristiche delle ali; e pterostigma. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come in entrambe le ali (anteriore e posteriore) di questa specie i triangoli presentino l'angolo acuto orientato verso la punta dell'ala. 4. Spazio mediano. 5. Membranule (notare come la membranula dell'ala anteriore sia più piccola di quella posteriore). 6. Campo sub-nodale. 7. Spazio cubitale. Notare che pur trattandosi di un maschio, in questa specie l'ala posteriore manca sia dell'angolo anale che del triangolo anale, mostrandosi in quella zona arrotondata. Continuiamo lo studio delle ali con un'altra specie: un maschio di Cordulegaster boltonii (Donovan, 1807) [famiglia Cordulegastridae]. Al solito: prima con foto "al naturale", seguita con foto e didascalie. Venature caratteristiche delle ali; e pterostigma. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come in entrambe le ali (anteriore e posteriore)di questa specie i triangoli presentino l'angolo acuto orientato verso la punta dell'ala. 4. Spazio mediano. 5. Membranule (notare come la membranula dell'ala anteriore sia più piccola di quella posteriore). 6. Triangolo anale. 7. Campo sub-nodale. 8. Angolo anale. Regioni particolari delle ali di un maschio di Onychogomphus forcipatus subsp. unguiculatus (Linnaeus, 1758) [famiglia Gomphidae]. 1. Campo discoidale. 2. Triangolo. Osservare come in entrambe le ali (anteriore e posteriore) di questa specie i triangoli presentino l'angolo acuto orientato verso la punta dell'ala. 3. Spazio mediano. 4. Sub-triangolo. 5. Triangolo anale. 6. Angolo anale. 7. Campo sub-nodale. 8. Campo anale. Analisi delle ali di una femmina (rinvenuta già morta al bordo di un torrente) di Somatochlora meridionalis (Nielsen, 1935) [famiglia Corduliidae]. Foto di Massimo Squarcini. Venature caratteristiche delle ali; e pterostigma. Viene anche posta l'attenzione su quella "Ultima venatura trasversa antenodale" dell'ala anteriore, osservando che è "completa". Torneremo in seguito su tale concetto e sulle venature trasverse antenodali; e vedremo come la loro osservazione può essere importante ai fini determinativi. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come in questa specie il triangolo dell'ala anteriore presenti l'angolo acuto rivolto verso la zona inferiore dell'ala; mentre il triangolo dell'ala posteriore presenta l'angolo acuto rivolto verso la punta dell'ala. 4. Spazio mediano. 5. Sub-triangolo 6. Campo sub-nodale. 7. Assenza di un triangolo anale dal contorno ben definito e marcato; caratteristica di tutte le femmine degli Anisotteri. 8. Spazio cubitale. Osservare la rotondità dell'ala posteriore, in assenza dell'angolo anale (e del triangolo anale); caratteristica di tutte le femmine degli Anisotteri. Analisi delle ali di una femmina di Libellula depressa (Linnaeus, 1758) [famiglia Libellulidae]. Venature caratteristiche delle ali; e pterostigma. Viene anche posta l'attenzione su quella "Ultima venatura trasversa antenodale" dell'ala anteriore, osservando che è "completa". Torneremo in seguito su tale concetto e sulle venature trasverse antenodali; e vedremo come la loro osservazione può essere importante ai fini determinativi. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come il triangolo dell'ala anteriore presenti l'angolo acuto rivolto verso la zona inferiore dell'ala; mentre il triangolo dell'ala posteriore presenta l'angolo acuto rivolto verso la punta dell'ala. 4. Spazio mediano. 5. Membranule. 6. Campo sub-nodale. Osservare la rotondità dell'ala posteriore, in assenza dell'angolo anale e del triangolo anale; caratteristica di tutte le femmine degli Anisotteri. Analisi delle ali di un maschio di Orthetrum albistylum (Sélys, 1848) [famiglia Libellulidae]. Venature caratteristiche delle ali; e pterostigma. Viene anche posta l'attenzione su quella "Ultima venatura trasversa antenodale" dell'ala anteriore, osservando che è "completa". Torneremo in seguito su tale concetto e sulle venature trasverse antenodali; e vedremo come la loro osservazione può essere importante ai fini determinativi. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come il triangolo dell'ala anteriore presenti l'angolo acuto rivolto verso la zona inferiore dell'ala; mentre il triangolo dell'ala posteriore presenta l'angolo acuto rivolto verso la punta dell'ala. 4. Spazio mediano. 5. Sub-triangolo. 6. Campo sub-nodale. 7. Spazio cubitale. Osservare la rotondità dell'ala posteriore, in assenza dell'angolo anale e del triangolo anale; caratteristica di tutte le specie (maschi e femmine) della famiglia Libellulidae. Analisi delle ali di un maschio di Orthetrum brunneum (Fonscolombe, 1317) [famiglia Libellulidae]. Venature caratteristiche delle ali; e pterostigma. Viene anche posta l'attenzione su quella "Ultima venatura trasversa antenodale" dell'ala anteriore, osservando che è "completa". Torneremo in seguito su tale concetto e sulle venature trasverse antenodali; e vedremo come la loro osservazione può essere importante ai fini determinativi. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come il triangolo dell'ala anteriore presenti l'angolo acuto rivolto verso la zona inferiore dell'ala; mentre il triangolo dell'ala posteriore presenta l'angolo acuto rivolto verso la punta dell'ala. 4. Spazio mediano. 5. Sub-triangolo. 6. Membranule. 7. Campo sub-nodale. 8. Spazio cubitale. Osservare la rotondità dell'ala posteriore, in assenza dell'angolo anale e del triangolo anale; caratteristica di tutte le specie (maschi e femmine) della famiglia Libellulidae. Analisi delle ali di una femmina (immatura) di Sympetrum meridionale (Sélys, 1841) [famiglia Libellulidae]. Venature caratteristiche delle ali; e pterostigma. Viene anche posta l'attenzione su quella "Ultima venatura trasversa antenodale" dell'ala anteriore osservando, stavolta, che è "incompleta". Torneremo in seguito su tale concetto e sulle venature trasverse antenodali; e vedremo come la loro osservazione può essere importante ai fini determinativi. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come il triangolo dell'ala anteriore presenti l'angolo acuto rivolto verso la zona inferiore dell'ala; mentre il triangolo dell'ala posteriore presenta l'angolo acuto rivolto verso la punta dell'ala. 4. Spazio mediano. 5. Sub-triangolo. 6. Membranule. 7. Campo sub-nodale. Osservare la rotondità dell'ala posteriore, in assenza dell'angolo anale e del triangolo anale; caratteristica di tutte le femmine degli Anisotteri. Analisi delle ali di un maschio di Sympetrum striolatum (Charpentier, 1840) [famiglia Libellulidae]. Venature caratteristiche delle ali. Viene anche posta l'attenzione su quella "Ultima venatura trasversa antenodale" dell'ala anteriore, osservando che è "incompleta". Torneremo in seguito su tale concetto e sulle venature trasverse antenodali; e vedremo come la loro osservazione può essere importante ai fini determinativi. Alcune regioni caratteristiche: 1. Campo discoidale. 2. Campo anale. 3. Triangolo. Osservare come il triangolo dell'ala anteriore presenti l'angolo acuto rivolto verso la zona inferiore dell'ala; mentre il triangolo dell'ala posteriore presenta l'angolo acuto rivolto verso la punta dell'ala. 4. Spazio mediano. 5. Membranula. 6. Sub-triangolo. 7. Campo sub-nodale. Osservare la rotondità dell'ala posteriore, in assenza dell'angolo anale e del triangolo anale; caratteristica di tutte le specie (maschi e femmine) della famiglia Libellulidae. Breve analisi delle ali di un maschio di Trithemis annulata (Palisot de Beauvois, 1807) [famiglia Libellulidae]. 1. Campo discoidale. 2. Triangolo. Osservare come il triangolo dell'ala anteriore presenti l'angolo acuto rivolto verso la zona inferiore dell'ala; mentre il triangolo dell'ala posteriore presenta l'angolo acuto rivolto verso la punta dell'ala. 3. Sub-triangolo. 4. Campo sub-nodale. 5. Ultima venatura trasversa antenodale dell'ala anteriore, incompleta. Torneremo in seguito su tale concetto e sulle venature trasverse antenodali; e vedremo come la loro osservazione può essere importante ai fini determinativi. Analisi dell'ala anteriore (vista da sotto) di una femmina di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]. Qui di seguito verranno proposti alcuni esempi per avere un'idea su quali indicazioni diagnostiche può fornire l'analisi e lo studio delle ali, soprattutto nei riguardi degli Anisotteri..jpg.313b9748bd308ac5b114d6c0a33803b4.jpg)
_-MESSA.jpg.3fee2cd062c53e93f5aadf34dfe7ccfc.jpg)
-MESSA.jpg.359c975cfd6523efd0ca26db91acdc83.jpg)
_MESSA.jpg.cc80ff7bfa49519eb78f5e69585d0e49.jpg)
---MESSA.jpg.d4a6ca4a48b790e31c84232d07e0ec3e.jpg)
---MESSA.jpg.d62fcb6dd9c63a98b48640b0a7c9f061.jpg)
---MESSA.jpg.edd85ec5fe340b76946fd036d49d6669.jpg)
---MESSA.jpg.5086a840f79bbeb0f160ea053d8e82a2.jpg)
.jpg.0a2f84a392bfd67fc8e5274709fb6f91.jpg)
.jpg.c595d738f889948a8bd1b988649f3cd5.jpg)
_MESSA.jpg.e1af87c78690033af02ed193701779c8.jpg)
---MESSA.jpg.d79db475a523dd9c98f2228fa9d7fb1b.jpg)

--MESSA.jpg.24727695861ed3cf60c2a28451ea6708.jpg)
---MESSA.jpg.4e820d83f49485db2f5a4a71ace5e561.jpg)
_MESSA.jpg.95dc06ab8e57ae0a2274f6ba3cd2aa4a.jpg)


_MESSA.jpg.6097cb6e77f6a626872a39a8de4e7b35.jpg)
_MESSA.jpg.cd8fe55ed4edb31241b18eb63449b1ee.jpg)


---Copia.jpg.9ca65188c29f453831bb685ed9f7d900.jpg)
-
ODONATA_ 03 - Sottordini dell’ordine Odonata, in ambito italiano
Alessandro F ha postato un topic nell'area Odonata
3_ Sottordini dell’ordine Odonata, in ambito italiano Come già esposto altrove, in questi appunti verrà mantenuta la suddivisione “classica” dell’ordine Odonata in 3 sottordini: Zygoptera (Sélys, 1854), Anisoptera (Sélys, 1854) e Anisozygoptera (Handlirsch, 1906); di cui soltanto i primi due riguardano la fauna europea, essendo gli Anisozygoptera esclusivi del continente asiatico e rappresentati da pochissime specie. Vedremo qui, a grandi linee, quali sono le caratteristiche di tali sottordini e quali sono le famiglie che li rappresentano. In alto: maschio immaturo di Trithemis annulata (Palisot de Beauvois, 1807) [sottordine Anisoptera; famiglia Libellulidae]; in basso: copula di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae] In alto: tandem in volo di Coenagrion scitulum (Rambur, 1842) [sottordine Zygoptera; famiglia Coenagrionidae]; in basso: femmina in ovideposizione di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae] Esempio di rappresentanti maschili dei due sottordini dell'ordine Odonata. A sinistra Calopteryx splendens (sottordine Zygoptera); a destra Onychogomphus forcipatus (sottordine Anisoptera). Pur avendo dimensioni simili in lunghezza (essendo i Calopterigidi tra gli Zigotteri più grandi e gli Onychogomphus tra gli Anisotteri più piccoli) la differenza che salta immediatamente agli occhi è il modo con cui vengono tenute le ali a riposo. Anche l'osservazione degli occhi non deve trarre in inganno: distanziati abbondantemente e relativamente piccoli tra gli Zigotteri; decisamente più grandi e o mai distanziati o un poco distanziati unicamente nelle specie della famiglia Gomphidae come, appunto, nel caso degli Onychogomphus. Femmina in ovideposizione di Anax imperator [sottordine Anisoptera; famiglia Aeshnidae]; mentre le vola attorno un maschio di Coenagrion puella [sottordine Zygoptera; famiglia Coenagrionidae] 3.1_ Zygoptera (Sélys, 1854) Questo taxon fu istituito nel 1854 dallo zoologo e politico belga Michel Edmond de Sélys Longchamps (1813–1900). L’etimologia risale al greco ζυγόν (zygòn) = giogo, unione, strumento che serve per unire; e al greco πτερόν = (pteròn) ala; quindi col significato di “con le ali unite”; in riferimento a quelle specie che, a riposo, tengono le ali riunite e disposte longitudinalmente al di sopra dell’addome. Considerando gli esemplari adulti (imago), le specie appartenenti a questo sottordine sono caratterizzate dall’avere le ali anteriori e posteriori simili tra di loro, sia nella forma che nelle venature; hanno occhi composti non eccessivamente grandi ma ben separati e sporgenti ai due lati della testa: in linea di massima la distanza che separa gli occhi è nettamente maggiore del diametro di un singolo occhio. Hanno dimensioni da piccole a medie; per le specie reperibili in Italia si può osservare una lunghezza variabile dai 25 mm per le specie più piccole ai 50 mm per le specie più grandi; con apertura alare che, a seconda della specie, varia tra 25 e 70 mm. Quando sono fermi e a riposo gli Zigotteri tengono le quattro ali più o meno tutte ravvicinate come le pagine chiuse di un libro e disposte longitudinalmente al di sopra dell’addome, in modo che non sempre è immediato rendersi conto del loro numero complessivo. Fanno eccezione le specie di alcuni generi della famiglia Lestidae che, a riposo, tengono invece le ali un poco aperte diagonalmente, in modo che è possibile distinguerle tutt’e quattro. Il volo degli Zigotteri è relativamente debole e lento (quanto meno se confrontato a quello degli Anisotteri) e in alcune specie può ricordare quello delle farfalle. Per le loro dimensioni e per la loro struttura esile e longilinea, con un addome sempre snello e allungato rispetto al torace, sono chiamati popolarmente "damigelle" o "donzelle", in ciò richiamando anche i nomi popolari riscontrabili in altre lingue come Demoiselles (signorine) in francese, Jungfer (zitella, signorina) in tedesco, Wasserjungfer (signorina acquatica) in tedesco, Damselfly (damigella con le ali) in inglese, ecc. Le femmine presentano sempre un ovopositore, più o meno evidente ed esposto, collocato sotto agli ultimi segmenti addominali. Attualmente (anno 2023; a meno di migrazioni, di periodiche assenze o di eventuali estinzioni), nel territorio italiano, il sottordine Zygoptera è rappresentato da 33 specie [o 32, secondo alcune fonti che considerano Coenagrion castellani (Roberts, 1948) come sottospecie di Coenagrion mercuriale (Charpentier, 1840)], raggruppate in 12 generi e 4 famiglie: # famiglia Calopterygidae rappresentata dal solo genere Calopteryx; # famiglia Coenagrionidae rappresentata dai generi Ceriagrion, Coenagrion, Enallagma, Erythromma, Ischnura, Nehalennia e Pyrrhosoma; # famiglia Lestidae rappresentata dai generi Chalcolestes, Lestes e Sympecma; # famiglia Platycnemididae rappresentata dal solo genere Platycnemis. Maschio di Chalcolestes viridis (Vander Linden, 1825) [famiglia Lestidae]. Osservare, con l'esemplare a riposo, le 4 ali semiaperte. Maschio di Platycnemis pennipes (Pallas, 1771) [famiglia Platycnemididae]. Stavolta, con l'esemplare a riposo, le ali sono molto ravvicinate e se non fosse per la presenza dei 4 pterostigma marroni non potremmo neanche individuarne il numero. Sulla destra dell'addome vengono riunite le ali anteriore e posteriore di destra; idem per la parte sinistra dell'addome. Come risulta evidente dalla foto seguente che ritrae ancora un maschio di Platycnemis pennipes. Si osserva anche che, a riposo, l’ala anteriore si colloca tra addome e ala posteriore. Maschio di Calopteryx virgo (Linnaeus, 1758) [famiglia Calopterygidae]. Nei maschi delle Calopterygidae sono assenti i pterostigma sulle ali; quindi, da questa foto, sarebbe impossibile stabilirne il numero essendo tutt'e 4 colorate e ravvicinate. Maschio di Pyrrhosoma nymphula (Sulzer, 1776) [famiglia Coenagrionidae]. Femmina di Calopteryx splendens (Harris, 1780) [famiglia Calopterygidae]; le femmine di questa famiglia mostrano su ciascuna delle 4 ali un "falso pterostigma" chiaro, composto da più celle e non da una singola cella (vedi foto qui sotto). Evidente anche l'ovodepositore sotto gli ultimi segmenti dell'addome. Femmina di Platycnemis pennipes (Pallas, 1771) [famiglia Platycnemididae]; evidente l'ovodepositore sotto gli ultimi segmenti dell'addome. Maschio di Ceriagrion tenellum (de Villers, 1789) [famiglia Coenagrionidae]. Foto di Massimo Squarcini. Maschio di Lestes virens (Charpentier, 1825) [famiglia Lestidae]. Osservare, con l'esemplare a riposo, le 4 ali semiaperte. Due Zigotteri a confronto: sulla sinistra una femmina di Calopteryx splendens (Harris, 1780) [famiglia Calopterygidae]; sulla destra un maschio di Platycnemis pennipes (Pallas, 1771) [famiglia Platycnemididae]. Tra tutti gli Zigotteri reperibili in Europa, gli esemplari della famiglia Calopterygidae sono quelli di dimensioni maggiori. Femmina di Sympecma fusca (Vander Linden, 1820) [sottordine Zygoptera; famiglia Lestidae]; evidente l'ovodepositore sotto gli ultimi segmenti dell'addome. Nel genere Sympecma le ali a riposo vengono tenute tutt'e 4 riunite proprio assieme in quanto vengono disposte da una unica parte dell'addome e non 2 per parte. Come risulta evidente nella foto precedente con le 4 ali disposte sulla destra dell'addome. Nella foto seguente che raffigura un maschio sempre di Sympecma fusca le 4 ali sono tenute sulla sinistra dell'addome. Femmina (nella “forma fosca”) di Enallagma cyathigerum (Charpentier, 1840) [famiglia Coenagrionidae]; evidente l'ovopositore sotto gli ultimi segmenti dell'addome. Femmine di Chalcolestes viridis (Vander Linden, 1825) [famiglia Lestidae]. Maschio di Erythromma viridulum (Charpentier, 1840) [famiglia Coenagrionidae]. Maschio di Calopteryx haemorrhoidalis (Vander Linden, 1825) [famiglia Calopterygidae]. Maschio di Lestes barbarus (Fabricius, 1798) [famiglia Lestidae]. Maschio di Ischnura elegans (Vander Linden, 1820) [famiglia Coenagrionidae]. Femmina di Coenagrion castellani (Roberts, 1948) [famiglia Coenagrionidae]. Maschio di Coenagrion castellani (Roberts, 1948) [famiglia Coenagrionidae]. 3.2_ Anisoptera (Sélys, 1854) Conosciuti in Italia anche col nome popolare di “libellule”, da non confondere tuttavia col termine scientifico Libellula (indicante il genere Libellula, Linnaeus 1758); talvolta sono indicati col nome di “dragoni”, termine preso in prestito dal gergo popolare inglese (Dragonfly). Questo taxon fu istituito nel 1854 dallo zoologo e politico belga Michel Edmond de Sélys Longchamps (1813–1900). L’etimologia risale al greco ἄνισος (ànisos) = diseguali [termine composto dalla particella negativa greca ἄν (an) = non, contrario; combinata con l’aggettivo greco ἴσος = (ìsos) uguale, medesimo], e al greco πτερόν (pteròn) = ala; quindi col significato di “con le ali diseguali”. Il termine latino-scientifico Anisoptera fu coniato nel 1829 dall’entomologo inglese James Francis Stephens (1792–1852) ma in relazione a un altro raggruppamento di insetti; venne infatti indicato con tale termine un insieme di insetti facenti parte degli Efemerotteri (ordine Ephemeroptera; insetti conosciuti anche col nome popolare di “effimere” e tassonomicamente imparentati con le libellule visto che, come già osservato in altra sede, Ephemeroptera e Odonata sono gli unici ordini inseriti nella sottocoorte Palaeoptera) e che mostravano il paio di ali anteriori grandi e di forma triangolare, decisamente diverso dal paio di ali posteriori più piccole e rotondeggianti. Successivamente, nel 1854, il Sélys Longchamps utilizzò il termine precedentemente coniato dallo Stephens, chiamando Anisoptera l’altro grande sottordine degli Odonata in riferimento alla diversità riscontrabile tra le ali. Considerando gli esemplari adulti (imago), le specie appartenenti a questo sottordine sono infatti caratterizzate dall’avere le ali anteriori e posteriori diverse tra di loro, sia nelle dimensioni (con le ali posteriori più grandi di quelle anteriori) sia nella venulazione. Altro modo per capire che siamo di fronte a esemplari facenti parte degli Anisotteri è quello di osservarli (se possibile) quando sono posati e a riposo: infatti, come già accennato, tengono le ali ben aperte, tutt’e quattro visibili e tenute più o meno perpendicolari rispetto all’asse testa-torace-addome. Hanno due occhi composti molto grandi, evidenti e prominenti a tal punto che, in molte specie e a una prima osservazione superficiale, sembra che la loro testa sia formata solo da questi due organi. Gli occhi sono inoltre ravvicinati tra di loro e, in base alla famiglia di appartenenza, si toccano più o meno estesamente lungo il loro lato interno, tranne che nelle specie della famiglia Gomphidae in cui gli occhi sono ben separati l’uno dall’altro ma comunque sempre a distanza minore del diametro di un singolo occhio. Il volo degli Anisotteri è veloce e potente, sempre ben coordinato e mai simile a quello delle farfalle (come invece può accadere per alcune specie di Zigotteri). A seconda delle specie, gli Anisotteri hanno dimensioni da medie a medio-grandi fino a decisamente grandi; per le specie reperibili in Italia si può osservare una lunghezza che varia da 30-35 mm fino a 80-90 mm; con apertura alare variabile tra 45-50 mm e 110-115 mm. Fa eccezione la Brachythemis impartita che rappresenta il più piccolo anisottero reperibile in Italia (soprattutto in Sicilia e Sardegna) e che può misurare, al minimo, 25 mm di lunghezza e 40 mm di apertura alare. L’addome si presenta assai più robusto e appariscente rispetto a quello degli Zigotteri. Solo le femmine della famiglia Aeshnidae hanno un ovopositore evidente. Attualmente (anno 2023; a meno di migrazioni, di periodiche assenze o di eventuali estinzioni), nel territorio italiano, il sottordine Anisoptera è rappresentato da 63 specie raggruppate in 26 generi e 6 famiglie: # famiglia Aeshnidae rappresentata dai generi Aeshna, Anax, Boyeria e Brachytron; # famiglia Cordulegastridae rappresentata dal solo genere Cordulegaster; # famiglia Corduliidae rappresentata dai generi Cordulia, Somatochlora; di tale famiglia fa parte anche il genere Epitheca che, un tempo, era rappresentato in Italia dalla specie Epitheca bimaculata. Tuttavia da più di un secolo non ci sono più segnalazioni di tale specie che è quindi da considerarsi assente/estinta nel nostro territorio; # famiglia Synthemistidae; in Europa (e in Italia) è rappresentata dal solo genere Oxygastra; genere che, a livello mondiale, è monospecifico contemplando la sola specie Oxygastra curtisii (Dale, 1834). Questa famiglia contempla soprattutto specie e generi tipici dell'Australia o della Nuova Guinea; qualche autore la considera ancora come una semplice sottofamiglia della famiglia Corduliidae. # famiglia Gomphidae rappresentata dai generi Gomphus, Lindenia, Onychogomphus, Ophiogomphus, Paragomphus e Stylurus; # famiglia Libellulidae rappresentata dai generi Brachythemis, Crocothemis, Diplachodes, Leucorrhinia, Libellula, Orthetrum, Pantala, Selysiothemis, Sympetrum, Tramea, Trithemis e Zygonyx. Maschi di Aeshna mixta (Latreille, 1805) [famiglia Aeshnidae]. Notare le 4 ali ben distinguibili perché, a riposo, aperte come le pagine di un libro; perpendicolari rispetto all'addome. Femmina di Libellula depressa (Linnaeus, 1758) [famiglia Libellulidae]. Femmina di Orthetrum albistylum (Sélys, 1848) [famiglia Libellulidae]. Maschio di Cordulegaster boltonii (Donovan, 1807) [famiglia Cordulegastridae]. A sinistra un maschio di Orthetrum cancellatum (Linnaeus, 1758) [famiglia Libellulidae]; a destra un maschio di Sympetrum striolatum (Charpentier, 1840) [famiglia Libellulidae]. Maschio di Onychogomphus forcipatus subsp. unguiculatus (Linnaeus, 1758) [famiglia Gomphidae]. A sinistra un maschio di Orthetrum cancellatum (Linnaeus, 1758) [famiglia Libellulidae]; a destra un maschio di Crocothemis erythraea (Brullé, 1832) [famiglia Libellulidae]. Femmina di Selysiothemis nigra (Vander Linden, 1825) [famiglia Libellulidae]. Foto di Enzo Orgera. Femmina di Somatochlora meridionalis (Nielsen, 1935) [famiglia Corduliidae]. Foto di Massimo Squarcini. Maschio di Libellula quadrimaculata (Linnaeus, 1758) [famiglia Libellulidae]. Femmina di Orthetrum cancellatum (Linnaeus, 1758) [famiglia Libellulidae]. Maschio di Sympetrum meridionale (Sélys, 1841) [famiglia Libellulidae]. Maschio di Anax imperator (Leach, 1815) [famiglia Aeshnidae]. Maschio di Crocothemis erythraea (Brullé, 1832) [famiglia Libellulidae]. A sinistra un maschio di Trithemis annulata (Palisot de Beauvois, 1807) [famiglia Libellulidae]; a destra un maschio di Orthetrum cancellatum (Linnaeus, 1758) [famiglia Libellulidae]. Femmina di Sympetrum fonscolombii (Sélys, 1840) [famiglia Libellulidae]. Maschio di Aeshna affinis (Vander Linden, 1820) [famiglia Aeshnidae]. Maschio di Orthetrum albistylum (Sélys, 1848) [famiglia Libellulidae]. A sinistra un maschio di Crocothemis erythraea (Brullé, 1832) [famiglia Libellulidae]; a destra un maschio di Trithemis annulata (Palisot de Beauvois, 1807) [famiglia Libellulidae]. Maschio di Aeshna isoceles (Müller, 1767) [famiglia Aeshnidae]. In "litigio" per accaparrarsi un posatoio ottimale. Un maschio di Orthetrum cancellatum (Linnaeus, 1758) [famiglia Libellulidae] cerca di scacciare un maschio di Crocothemis erythraea (Brullé, 1832) [famiglia Libellulidae] saltandogli addosso. A destra un maschio in volo di Crocothemis erythraea (Brullé, 1832) [famiglia Libellulidae]; a sinistra un maschio di Trithemis annulata (Palisot de Beauvois, 1807) [famiglia Libellulidae]. A destra un maschio di Crocothemis erythraea (Brullé, 1832) [famiglia Libellulidae]; nel centro un maschio in volo di Orthetrum albistylum (Sélys, 1848) [famiglia Libellulidae]; a sinistra un maschio di Trithemis annulata (Palisot de Beauvois, 1807) [famiglia Libellulidae]. In "litigio" per accaparrarsi un posatoio ottimale. Un maschio di Crocothemis erythraea (Brullé, 1832) [famiglia Libellulidae] cerca di scacciare un maschio di Orthetrum albistylum (Sélys, 1848) [famiglia Libellulidae] saltandogli addosso. 3.3_ Anisozygoptera (Handlirsch, 1906) Il nome di questo sottordine deriva da una combinazione dei nomi dei due sottordini Anisoptera e Zygoptera. In effetti le poche specie di questo sottordine hanno ali simili a quelle degli Zigotteri ma nel loro aspetto complessivo e nelle dimensioni ricordano meglio gli Anisotteri. Come già accennato più sopra tali specie non riguardano la fauna europea.

.jpg.ecbad928202d363ba6756d79d0ea6125.jpg)

.jpg.4dc01f6e4e5bb3e8f9cacdb660c4d0f4.jpg)
.jpg.2cdff849ba42bdedb625f0a0746c7cbb.jpg)
.jpg.da402926de4a5ac4c784a8f22f866ca1.jpg)
.jpg.e4f3f7382b3420e5c8e7914d023b3bec.jpg)
.jpg.52441bb107b6ec6d5874c66cb0c7b994.jpg)
.jpg.eaedf48fa22e5fd81a6d948327e1b6e1.jpg)
.jpg.7901778ef978b15384a1887bdcf5f96b.jpg)
.jpg.e8499d21b61ced38c0755680cd55ba0a.jpg)
.jpg.c18076f84dfe23108d56495382c12ce8.jpg)
.jpg.441b8ae0e53b60340388376bd1270d1f.jpg)
.jpg.2676640941c6a9bc4777f26973cb92e4.jpg)
.jpg.500a8deaa11da7b63268094b50a8123f.jpg)










.jpg.fcc09991e51c6cf1ff4e827db1038195.jpg)
.jpg.2be14af76f15324421734a8cbbc78a6d.jpg)
.jpg.ccfc64a735c7a61d4e92340e19515bcf.jpg)
.jpg.433b4804f930ce3ee79d2ec6dc13af2d.jpg)
.jpg.c65b254199378271de291fad734f1e4a.jpg)

.jpg.1d662feaa7f2281b366283e4459b0c4d.jpg)

.jpg.cda1cf56820ed6ef7fab6d6efcde2b83.jpg)
.jpg.682a5edeaf569be24437046fe40ee0aa.jpg)

-(2).jpg.8e60bd6bb80f8cec0d2588e747382137.jpg)











-
ODONATA_ 02 - Ordine Odonata (Fabricius, 1793)_ Caratteri generali
Alessandro F ha risposto alla discussione di Alessandro F in Odonata
N.B. Per uno studio e una analisi approfondita delle ali e delle loro venature, vedere qui. 2.5_ L'addome L’addome si presenta sempre particolarmente allungato rispetto alla compattezza del torace. È connesso (sessile) direttamente col torace, senza presentare alcun peduncolo di collegamento come accade, per esempio, nelle vespe. Nell’addome sono riconoscibili 10 segmenti (o uriti) ben differenziati, con un undicesimo segmento finale talvolta appena abbozzato ma spesso non ben percepibile o assente e quindi non considerato nelle varie descrizioni. Nelle osservazioni occorre prestare attenzione al conteggio del 1° segmento addominale: spesso non viene preso in considerazione perché abbastanza esiguo rispetto agli altri e/o nascosto sotto la parte posteriore del torace. Anche i disegni, soprattutto se scuri, presenti sull'addome possono creare difficoltà nel conteggio. L'addome termina con particolari appendici (dette anche cerci) di numero variabile: sempre soltanto 2 nelle femmine, 3 nei maschi degli Anisotteri, 4 nei maschi degli Zigotteri. La loro funzione verrà esaminata in altra sede, nelle osservazioni relative all'accoppiamento. Nella parte inferiore dell'addome sono visibili gli organi sessuali maschili e femminili, nonché, in tutte le femmine degli Zigotteri e di alcune famiglie di Anisotteri, il cosiddetto apparato ovopositore. La loro osservazione, assieme a quella delle appendici addominali, consente di distinguere facilmente i maschi dalle femmine; per maggiori dettagli, vedere qui. I 10 segmenti addominali di una femmina di Platycnemis pennipes (Pallas, 1771) [sottordine Zygoptera; famiglia Platycnemididae]. Nelle visioni dorsali il 1° segmento non è mai ben visibile. Segmenti addominali di una femmina di Platycnemis pennipes (Pallas, 1771) [sottordine Zygoptera; famiglia Platycnemididae]. Sotto a 8° e 9° segmento sono ben visibili l'ovopositore e la vulva. Segmenti addominali di un maschio di Platycnemis pennipes (Pallas, 1771) [sottordine Zygoptera; famiglia Platycnemididae]. Nelle visioni dorsali il 1° segmento non è mai ben visibile. Segmenti addominali di un maschio di Platycnemis pennipes (Pallas, 1771) [sottordine Zygoptera; famiglia Platycnemididae]. In foto con questa angolazione è facile osservare la presenza dell'apparato copulatore maschile (una sorta di pene) al di sotto del 2° segmento (nelle femmine tale zona rimane completamente piatta) e, sotto al 9° segmento, lo sbocco del canale seminale. Segmenti addominali di un maschio di Coenagrion scitulum (Rambur, 1842) [sottordine Zygoptera; famiglia Coenagrionidae]. Visibile, sotto al 2° segmento, l'apparato copulatore. Segmenti addominali di un maschio di Calopteryx haemorrhoidalis (Vander Linden, 1825) [sottordine Zygoptera; famiglia Calopterygidae]. Visibile, sotto al 2° segmento, l'apparato copulatore. Segmenti addominali in una femmina di Enallagma cyathigerum (Charpentier, 1840) [sottordine Zygoptera; famiglia Coenagrionidae]. Sotto a 8° e 9° segmento sono ben visibili l'ovopositore e la vulva. Segmenti addominali in una femmina di Erythromma lindenii (Selys, 1840) [sottordine Zygoptera; famiglia Coenagrionidae]. Segmenti addominali in una femmina di Ischnura pumilio (Charpentier, 1825) [sottordine Zygoptera; famiglia Coenagrionidae]. Sotto a 8° e 9° segmento sono ben visibili l'ovopositore e la vulva. Segmenti addominali di un maschio di Pyrrhosoma nymphula (Sulzer, 1776) [sottordine Zygoptera; famiglia Coenagrionidae]. Nelle visioni dorsali il 1° segmento non è mai ben visibile. Segmenti addominali in una femmina di Pyrrhosoma nymphula (Sulzer, 1776) [sottordine Zygoptera; famiglia Coenagrionidae]. Sotto a 8° e 9° segmento sono ben visibili l'ovopositore e la vulva. Segmenti addominali di una femmina di Lestes virens (Charpentier, 1825) [sottordine Zygoptera; famiglia Lestidae]. Sotto a 8° e 9° segmento sono ben visibili l'ovopositore e la vulva. Segmenti addominali in un maschio di Sympecma fusca (Vander Linden, 1820) [sottordine Zygoptera; famiglia Lestidae]. Segmenti addominali in un maschio di Aeshna affinis (Vander Linden, 1820) [sottordine Anisoptera; famiglia Aeshnidae]. La loro individuazione può essere resa complicata dai disegni addominali. Al di sotto del 2° segmento si intravede l'apparato copulatore maschile. Notare anche la disposizione delle zampe durante il volo. Negli Anisotteri le zampe vengono disposte nel modo più aereodinamico possibile per facilitare la velocità e l'agilità di volo: il paio di zampe anteriori è ripiegato e tenuto nascosto in verticale tra il retro degli occhi e il torace; la altre due paia di zampe sono tenute ripiegate e raccolte a contatto con la parte inferiore del torace. Segmenti addominali in un maschio di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]. La loro individuazione può essere resa complicata dai disegni addominali. Segmenti addominali in un maschio di Aeshna mixta (Latreille, 1805) [sottordine Anisoptera; famiglia Aeshnidae]. La loro individuazione può essere resa complicata dai disegni addominali. Segmenti addominali in un maschio di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]. La loro individuazione può essere resa complicata dai disegni addominali. Segmenti addominali in un maschio di Cordulegaster boltonii (Donovan, 1807) [sottordine Anisoptera; famiglia Cordulegastridae]. La loro individuazione può essere resa complicata dai disegni addominali. Segmenti addominali in un maschio di Onychogomphus forcipatus subsp. unguiculatus (Linnaeus, 1758) [sottordine Anisoptera; famiglia Gomphidae]. La loro individuazione può essere resa complicata dai disegni addominali. Segmenti addominali in un maschio di Aeshna juncea (Linnaeus, 1758) [sottordine Anisoptera; famiglia Aeshnidae]. La loro individuazione può essere resa complicata dai disegni addominali. Notare anche la disposizione delle zampe durante il volo. Negli Anisotteri le zampe vengono disposte nel modo più aereodinamico possibile per facilitare la velocità e l'agilità di volo: il paio di zampe anteriori è ripiegato e tenuto nascosto in verticale tra il retro degli occhi e il torace; la altre due paia di zampe sono tenute ripiegate e raccolte a contatto con la parte inferiore del torace. Segmenti addominali in un maschio di Orthetrum coerulescens (Fabricius, 1798) [sottordine Anisoptera; famiglia Libellulidae]. Nelle visioni dorsali il 1° segmento non è mai ben visibile. Segmenti addominali in un maschio di Orthetrum cancellatum (Linnaeus, 1758) [sottordine Anisoptera; famiglia Libellulidae]. Segmenti addominali in un maschio immaturo di Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae]. La loro individuazione può essere resa complicata dai disegni addominali. Al di sotto del 2° segmento si intravede l'apparato copulatore maschile. Segmenti addominali in una femmina di Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae]. Nella zona ventrale fra 8° e 9° segmento è ben visibile la lamina vulvare. Segmenti addominali in un maschio immaturo di Sympetrum fonscolombii (Sélys, 1840) [sottordine Anisoptera; famiglia Libellulidae]. Al di sotto del 2° segmento si intravede l'apparato copulatore maschile. Segmenti addominali in una femmina di Trithemis annulata (Palisot de Beauvois, 1807) [sottordine Anisoptera; famiglia Libellulidae]. Segmenti addominali in una femmina di Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]. Per quanto riguarda gli Anisotteri potrà essere utile constatare, con opportune inquadrature, la presenza o meno sui fianchi del 2° segmento addominale di due piccole protuberanze, dette "orecchiette laterali". Ciò consentirà di circoscrivere la determinazione ad alcune famiglie o anche a particolari generi. Inoltre la loro presenza indica che siamo in presenza di un maschio; non vale però il viceversa visto che molte famiglie di Anisotteri non presentano mai orecchiette, né nei maschi né nelle femmine. Assenza di "orecchiette laterali" sui fianchi del 2° segmento addominale, in un maschio di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]. Presenza di evidenti "orecchiette laterali" sui fianchi del 2° segmento addominale, in un maschio di Onychogomphus forcipatus subsp. unguiculatus (Linnaeus, 1758) [sottordine Anisoptera; famiglia Gomphidae]. Presenza di "orecchiette laterali" sui fianchi del 2° segmento addominale (qui visibile solo quella sinistra), in un maschio di Aeshna mixta (Latreille, 1805) [sottordine Anisoptera; famiglia Aeshnidae]. Assenza di "orecchiette laterali" sui fianchi del 2° segmento addominale, in un maschio di Anax ephippiger (Burmeister, 1839) [sottordine Anisoptera; famiglia Aeshnidae]. Presenza di "orecchiette laterali" sui fianchi del 2° segmento addominale, in un maschio di Aeshna affinis (Vander Linden, 1820) [sottordine Anisoptera; famiglia Aeshnidae]. Assenza di "orecchiette laterali" sui fianchi del 2° segmento addominale, in una femmina di Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]. Assenza di "orecchiette laterali" sui fianchi del 2° segmento addominale, in un maschio di Sympetrum meridionale (Sélys, 1841) [sottordine Anisoptera; famiglia Libellulidae]. Presenza di "orecchiette laterali" sui fianchi del 2° segmento addominale (qui visibile solo quella sinistra), in un maschio di Aeshna mixta (Latreille, 1805) [sottordine Anisoptera; famiglia Aeshnidae]. Presenza di "orecchiette laterali" sui fianchi del 2° segmento addominale, in un maschio immaturo di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]..jpg.3ba6c71c62665c8982d487f453eb65aa.jpg)
.jpg.3c9d43d6a9dfaa8e036412a93f65f777.jpg)
.jpg.efa2d127e6dc52033d8a427dd76f23ae.jpg)
.jpg.8478569b1c922b94ffefb87102318538.jpg)
.jpg.86f9b4db4e0ce2be8a8dd17aa1fbf8a0.jpg)
.jpg.f37d9493375552f7754f536f38b77163.jpg)
.jpg.7f3aa2317970079d3b82ce44a255c33b.jpg)
.jpg.e4d6d6f0cbd1c9d1520afa1ff3e6cd05.jpg)
.jpg.5a7e639ef18896f74e9421f8db1f5ca2.jpg)
.jpg.2278eba3a2f7f53cd433ab240dba8e30.jpg)
.jpg.9bffc8c9d58fa964d506020d51c9042c.jpg)
.jpg.055ff3a64ba236faf5297d66af07d1b7.jpg)
.jpg.78e74e8f587e5570eb2c6e8ea72f8510.jpg)
.jpg.ee13153a1a3b37e8b7fcc67ed7818ba2.jpg)
.jpg.7c16529c7675da2d1c4617a3e875fe5e.jpg)

.jpg.f5825a8c813e842931597536b130744c.jpg)
.jpg.25203ef82c57213cf9517985fe932a06.jpg)
.jpg.8ab71b0f3f944640e72b69ffcb92aa3e.jpg)
.jpg.4225cb7f16c699f9b2cfb30f8941fbed.jpg)
.jpg.cfd04e41905c1da60048a887c51fccbd.jpg)
.jpg.e005b713ac5ad1db9ed2f1d116c826c1.jpg)
.jpg.b29beb8a2bf31c9d5de28deab01035cc.jpg)
.jpg.a06c545ccc73314d4b2dc8ac8c69d56d.jpg)
.jpg.0ac2343a6c320df8f3e1b22b2337bcfc.jpg)
.jpg.45820550d1addb6af06d39bfaffebf37.jpg)
.jpg.06844191159ce705b6b89775329dca0d.jpg)
.jpg.31952be2a2e7523bd12082c134f411cb.jpg)
.jpg.5fda8e15d912c20ff5849b45a1eb3533.jpg)
.jpg.acc2078bd733f60b81e8b5c618fbec3a.jpg)
.jpg.d0688999c3ff89f6166ec8f4cc9fb885.jpg)
.jpg.1702c48843919bc2b7ca2045e57111e8.jpg)
.jpg.c3817404faab167559e253edd926784a.jpg)
.jpg.cb1503b0d54cbb230db138c3718b8017.jpg)
.jpg.9ec0893132d35dc9b6d76d2495d38f8e.jpg)
.jpg.5e94335d60a3383d676abd1d11addf92.jpg)
-
ODONATA_ 02 - Ordine Odonata (Fabricius, 1793)_ Caratteri generali
Alessandro F ha risposto alla discussione di Alessandro F in Odonata
2.4_ Il torace e le zampe Il torace delle libellule, come per molti altri esapodi, è la sede degli organi preposti al moto. Al di sotto di esso vi sono 3 paia di zampe mentre al di sopra si aprono 2 paia di ali efficacemente strutturate e specializzate per il volo. Le zampe, in pratica, non vengono mai impiegate per la deambulazione visto che le libellule si spostano esclusivamente in volo; viceversa rappresentano strutture necessarie per ancorarsi e posizionarsi sui vari posatoi e sono a tutti gli effetti dei micidiali strumenti di caccia essendo munite di abbondanti e ispide setole indispensabili per afferrare e trattenere le prede catturate in volo; sono infatti definite zampe del tipo “raptatorio”. A grandi linee il torace può essere suddiviso in tre regioni più o meno fuse tra di loro: sul retro della testa e dopo il brevissimo collo si colloca il "protorace", di seguito il "mesotorace" e infine, direttamente a contatto con l’addome, il "metatorace". Ognuno di essi porta un paio di zampe, le anteriori al di sotto del protorace, le mediane sotto al mesotorace, le posteriori sotto al metatorace; mentre le ali sono presenti, un paio per ciascuno, solo su meso– e metatorace. Apparentemente curiosa è la disposizione di meso– e metatorace: rispetto a un ideale asse longitudinale “testa-addome”, essi sono orientati non perpendicolarmente ma in obliquo (negli Anisotteri) o quasi in parallelo (negli Zigotteri) in modo che le loro parti ventrali si trovino nettamente più avanti (verso la testa) rispetto alle parti dorsali. Ciò comporta una funzionalità che conferma, se ce ne fosse bisogno, come le libellule siano dei perfezionati cacciatori volanti: le zampe ben lunghe, articolate nelle loro componenti e posizionate più verso la testa rispetto alle ali, permettono alle libellule (che cacciano, ricordiamolo, esclusivamente in volo) di effettuare le catture senza sbilanciarsi o perdere il proprio “assetto di volo”. Le zone (o placche) dorsali vengono solitamente indicate tramite il suffisso “–noto”; così troviamo “pronoto”, “mesonoto” e “metanoto” rispettivamente nella parte dorsale di protorace, mesotorace e metatorace. Quindi, per quanto detto sopra, il mesonoto e il metanoto portano ciascuno un paio di ali. Tra questi nomi sarà però “pronoto” quello che sicuramente ricorre più frequentemente nella descrizione delle varie specie perché la sua osservazione rende più agevole la determinazione delle specie studiate, soprattutto per quanto riguarda gli Zigotteri e le loro femmine. Le parti ventrali, tramite il suffisso “–sterno”, sono indicate rispettivamente con prosterno, mesosterno e metasterno; sono tuttavia parti non rilevanti ai fini descrittivi e determinativi. Importanti da osservare sono anche le zone laterali (o pleure) soprattutto del meso– e del metatorace; infatti disegni, bande (o strie) che le percorrono, colori e suture, possono contribuire in maniera decisiva alla determinazione della specie studiata. Per quanto riguarda le suture se ne possono osservare tre principali. Una "sutura interpleurale" che separa le parti laterali di meso– e matatorace, tuttavia non sempre ben definita o marcata. Un’altra sutura, detta "sutura omerale", si trova internamente alla parte laterale del mesotorace, dividendo quest’ultimo in due zone (episterno mesotoracico al di sopra e epimero mesotoracico al di sotto); in alcune specie, nell’episterno mesotoracico si trova una banda colorata, detta "stria anteomerale", la cui grandezza o conformazione può essere indicativa ai fini diagnostici. Una ulteriore sutura, detta "sutura metapleurale", si trova internamente alla parte laterale del metatorace, dividendo quest’ultimo in due zone (episterno metatoracico al di sopra e epimero metatoracico al di sotto); nell’episterno metatoracico è facile osservare, soprattutto negli Anisotteri, un piccolo forellino: si tratta di uno "stigma", un’apertura, che permette alle libellule di respirare. Vi sono altri stigma respiratori distribuiti tra torace e addome ma quello metatoracico è il più evidente. Maschio immaturo di Sympetrum meridionale (Sélys, 1841) [sottordine Anisoptera; famiglia Libellulidae]. Notare anche le zampe ben munite di ispide setole; e la "obliquità" di meso- e metatorace rispetto a un ideale asse longitudinale "testa-addome". Femmina di Platycnemis pennipes (Pallas, 1771) [sottordine Zygoptera; famiglia Platycnemididae]. Notare anche le zampe ben munite di ispide setole; evidente anche l'orientamento di meso- e metatorace che sono quasi paralleli rispetto a un ideale asse longitudinale "testa-addome". Maschio immaturo di Sympetrum fonscolombii (Sélys, 1840) [sottordine Anisoptera; famiglia Libellulidae]. Maschio di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]. Giovane femmina di Coenagrion puella (Linnaeus, 1758) [sottordine Zygoptera; famiglia Coenagrionidae]; foto di Massimo Squarcini. Altre zone individuabili a grandi linee nel torace sono le strutture che sorreggono le zampe come i due infraepisterni meso– e matatoracico e le tre anche. Nelle zampe sono distinguibili, nell’ordine e a partire dall’anca: trocantere, femore, tibia e tarso; quest’ultimo termina con un pretarso munito di piccoli e brevi artigli uncinati. Maschio immaturo di Sympetrum fonscolombii (Sélys, 1840) [sottordine Anisoptera; famiglia Libellulidae]. Maschio di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]..jpg.49dc4451fbeef07443ea0df1f1dbd7f1.jpg)
.jpg.d67b2ef31e2bae1af4e9f56050dc3716.jpg)
---Copia.jpg.02b6ad6a4fc9bc6ef45c60d84418d22f.jpg)
---Copia.jpg.39fe47f436a3343abff05ebcc344347b.jpg)
---Copia.jpg.5edda58b5589846cc6662b3786012671.jpg)
---Copia---Copia.jpg.5afa3aab477aa3a19197de41544e6439.jpg)
---Copia.jpg.5e3ee6de8604b76fa626237f7982688f.jpg)
---Copia.jpg.d3585c2d777971152f386aab3ddec9b7.jpg)
---Copia.jpg.1a182b8ce641438e6f8f8400d8450c1f.jpg)

-
ODONATA_ 02 - Ordine Odonata (Fabricius, 1793)_ Caratteri generali
Alessandro F ha risposto alla discussione di Alessandro F in Odonata
In ogni caso la vista delle libellule è talmente perfezionata e funzionale che alcune specie tra le più grandi possono individuare un piccolo insetto come ad esempio una mosca, ferma o in volo, anche a qualche decina di metri di distanza. Sembra infatti, da varie osservazioni ed esperimenti, che i più grandi Anisotteri riescano a individuare una preda immobile di dimensioni simili a quella di una mosca domestica a 15-20 metri di distanza, mentre se la preda è in movimento la distanza può arrivare a 40-50 metri. In proporzione sarebbe come se una persona riuscisse ad individuare un pallone da calcio in movimento da oltre un chilometro di distanza. Sempre sulla testa, nella zona facciale superiore (o vertice facciale), si notano due piccole e brevi antenne filiformi che rappresentano, come per la maggior parte degli insetti, particolari sensori biochimici; vi sono poi tre "ocelli" (o occhi semplici) disposti a triangolo: con un singolo ocello anteriore (solitamente il più grande) e due retrostanti. Diversamente dagli ommatidi che formano gli occhi composti, gli ocelli non percepiscono le immagini ma sono sensibili all’intensità luminosa e alla luce polarizzata. Sulla testa ma sul retro degli occhi degli Anisotteri spesso è visibile una piccola regione triangolare detta, appunto, “triangolo occipitale” che in alcuni casi può essere utile osservare per ottenere qualche indicazione in più ai fini determinativi. Ocelli e antenne di un maschio di Pyrrhosoma nymphula (Sulzer, 1776) [sottordine Zygoptera; famiglia Coenagrionidae]. Femmina androcroma di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]. Femmina immatura nella "forma rufescens" di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]. Maschio di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]. Triangolo occipitale di un maschio di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]. Triangolo occipitale di un maschio immaturo di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]. Triangolo occipitale e ocelli di un maschio di Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]. Gli ocelli posteriori di un maschio di Onychogomphus forcipatus subsp. unguiculatus (Linnaeus, 1758) [sottordine Anisoptera; famiglia Gomphidae]. Ocelli e antenne di una femmina di Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]. Ocelli di una femmina immatura di Orthetrum cancellatum (Linnaeus, 1758) [sottordine Anisoptera; famiglia Libellulidae]. Nella zona centrale della faccia, sotto al vertice, si nota una regione piuttosto evidente detta fronte. Al di sotto della fronte si trova il cosiddetto clipeo suddiviso in post-clipeo (al di sopra) e ante-clipeo. Sotto al clipeo c’è un efficace apparato masticatore formato da un labbro superiore (o labrum), due mandibole, due mascelle, e un labbro inferiore (o labium). Le due mandibole rimangono per lo più nascoste dal labbro superiore; le mandibole sovrastano le due mascelle, mobili e poste a loro volta nella zona inferiore della faccia al di sopra del labbro inferiore. La collaborazione coordinata tra mandibole e mascelle, tutte munite di una sorta di denti taglienti e controllate da una potente muscolatura, fa sì che l’apparato masticatore delle libellule sia così perfezionato da essere in grado di tranciare anche la corazza di piccoli coleotteri o di spezzare in due una mosca anche con un solo morso. E ciò vale sia per le libellule più grandi che per le più piccole. Maschio di Orthetrum coerulescens (Fabricius, 1798) [sottordine Anisoptera; famiglia Libellulidae]. Maschio di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]; fotografato mentre sta detergendosi gli occhi con le zampe anteriori grazie a una ampia rotazione della testa attorno al collo. Rotazione che ha permesso di vederlo "a bocca aperta". Maschio di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]. Femmina di Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]. Femmina di Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae]. Maschio di Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae]. Stranamente questo maschio ha compiuto un'operazione di volo sbagliata, andando a infilarsi e a intrappolarsi nelle maglie di una rete che sovrasta una vasca di raccolta di acqua. Vista la proverbiale abilità di volo delle libellule (soprattutto per gli Anisotteri), la manovra sbagliata potrebbe essere una conseguenza del frequente "litigio" aereo che i maschi compiono tra di loro per il controllo territoriale, compiendo voli funambolici e scontrandosi anche a grande velocità. L'esemplare è stato delicatamente liberato dalle maglie della rete, non senza aver prima approfittato della sua involontaria posizione per fotografarne l'apparato masticatore. Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]. Qui di seguito alcune "scene di caccia" e di conseguente "degustazione" da parte di varie libellule. Calopteryx haemorrhoidalis (Vander Linden, 1825) [sottordine Zygoptera; famiglia Calopterygidae]; maschio che mangia un insetto alato (scartandone le ali). Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]; femmina immatura (nella "forma violacea") che sta mangiando. Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]; maschio che mangia una mosca dopo averla spezzata in due.. Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]; maschio in caccia. Foto di Massimo Squarcini. Interessante osservare le zampe della libellula. Durante il volo perlustrativo gli Anisotteri tengono le zampe aderenti al corpo per favorire l'assetto aereodinamico: il paio di zampe anteriori ripiegate e raccolte in verticale tra torace e retro degli occhi, le altre due paia di zampe ripiegate e raccolte sotto al torace. Ma, come in questa foto nell'attimo che precede la cattura, le due paia di zampe mediane e posteriori lasciano la posizione "a riposo" e si preparano a ghermire la preda. La quale preda sembra essersi accorta del pericolo incombente e si dispone in un assetto difensivo, verticale e inconsueto... ma sicuramente per essa non ci sarà scampo. Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]; femmina immatura, nella forma "rufescens", che sta mangiando una femmina di Euaresta aequalis, mentre un maschio della stessa specie della preda si è avvicinato... Ischnura pumilio (Charpentier, 1825) [sottordine Zygoptera; famiglia Coenagrionidae]; una femmina ha catturato una "cicalina verde" (Cicadella viridis). Il malcapitato insetto che, come stazza, è quanto meno paragonabile al torace della damigella sembra non aver scampo. Ma la damigella ha sbagliato l'attacco: invece di afferrare la cicalina per il ventre la sta addentando sul dorso, evidentemente più coriaceo e protetto, e non riesce a far presa con i denti nelle parti vitali della preda, pur provando a mordere in vari punti... Orthetrum cancellatum (Linnaeus, 1758) [sottordine Anisoptera; famiglia Libellulidae]; una femmina vecchia sta mangiando una mosca, rigirandosela in bocca senza alcun aiuto delle zampe. Orthetrum coerulescens (Fabricius, 1798) [sottordine Anisoptera; famiglia Libellulidae]; una femmina vecchia sta mangiando una falena. In queste foto di Massimo Squarcini, scattate a pochi millisecondi una dall'altra, una sequenza dell'attacco nei confronti di un'ape da parte di un maschio di Orthetrum cancellatum (Linnaeus, 1758) [sottordine Anisoptera; famiglia Libellulidae] e di un maschio (in secondo piano e sfocato) di Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]. Nella seconda foto già si vedono le zampe mediane dell'Orthetrum cancellatum che si stanno per distendere così come è evidente il suo repentino cambio di direzione dovuto all'improvviso cambio di "assetto di volo" da parte dell'ape che, vedendosi attaccata da due parti, prova ad eludere l'imminente pericolo ribaltandosi sul dorso... (foto qui sotto) Anche l'osservazione della parte posteriore della testa può fornire indicazioni utili per il riconoscimento di alcune specie. In particolar modo in relazione alla presenza/assenza delle cosiddette "macchie post-oculari" o di altre peculiarità. Macchie post-oculari tipicamente dentellate inferiormente in una femmina di Coenagrion puella (Linnaeus, 1758) [sottordine Zygoptera; famiglia Coenagrionidae]. Macchie post-oculari giallognole in un maschio di Aeshna juncea (Linnaeus, 1758) [sottordine Anisoptera; famiglia Aeshnidae]; uno dei caratteri che permette di non confonderla con la congenere e, in Italia assai rara, Aeshna subartica (Walker, 1908). Assenza di macchie post-oculari in un maschio di Pyrrhosoma nymphula (Sulzer, 1776) [sottordine Zygoptera; famiglia Coenagrionidae]. Maschie post-oculari in un maschio di Enallagma cyathigerum (Charpentier, 1840) [sottordine Zygoptera; famiglia Coenagrionidae].
.jpg.e0d860552689b3b12b5d93721b91f0d4.jpg)
.jpg.800075cc7f6c44af27c602cb98de2432.jpg)
.jpg.837ea67ff416a5e6c57aa9e5b46b6360.jpg)
.jpg.e14a5028bd105d3618f7df5d87325fd9.jpg)




.jpg.952da7dcc232ffadbdf957a9c2f1e2fb.jpg)
.jpg.7b2c543212ab58a5a27b017a8057f8ad.jpg)
.jpg.2722f104bb5335602e0ec7ca78a717a5.jpg)
.jpg.1522635ef2811d95c712ecaa1086416b.jpg)
.jpg.a0d4dea798f946ea59b485a10e361422.jpg)
.jpg.5b263a0cd466b6803927d2d19592579b.jpg)
.jpg.e006ac7016283e347ac65eb6e2f8b943.jpg)
.jpg.24a07346b6f118216aa3a1d5a4882a2a.jpg)

.jpg.d944693d7f5b0e1d500bae49e20562aa.jpg)
.jpg.0b124e8f6b79e6cd22e6476618d321a7.jpg)
.jpg.0a8c1e82d8ed83eaf26eac78d7e2f991.jpg)
.jpg.8a34c02876c86a28d92906409d68ef01.jpg)
.jpg.8265bb0b0ef78b5aa05d4c285f43f7a9.jpg)
.jpg.141fe474ff44ad6590a2ce8a3fc67866.jpg)
.jpg.f88d606ce72319924afc1111b08f5fd4.jpg)
---Copia.jpg.7d42ded7465863b67fca8560531957f0.jpg)
.jpg.b250402b20e0fa41f794c68ea0505adf.jpg)




.jpg.2ac954cdeb9fd2e922851527c090cb72.jpg)
.jpg.2fe3d040bff5075c9e5fffb90930f65b.jpg)
-
ODONATA_ 02 - Ordine Odonata (Fabricius, 1793)_ Caratteri generali
Alessandro F ha postato un topic nell'area Odonata
2_ Ordine Odonata (Fabricius, 1793)_ Caratteri generali 2.1_ Cenni tassonomici ed etimologia dei termini Libellula e Odonata Il nome Libellula ha una gestazione assai particolare. Semplificando molto ci si potrebbe accontentare di una delle tre seguenti versioni: 1) proviene dal latino libellus = piccolo libro, in quanto le libellule in volo tengono le ali come le pagine aperte di un libretto; 2) proviene dal latino libellus = piccolo libro, in quanto le libellule più grandi (cioè gli odierni Anisotteri) quando sono ferme a riposo si dispongono con le ali ben aperte e simili alle pagine spalancate di un libretto; 3) proviene dal latino libella, a sua volta diminutivo del latino libra = bilancia a due bracci, in quanto nel volo le libellule tengono le ali orizzontali. In realtà il percorso etimologico del termine Libellula è diverso, più complesso e, per certi versi, sorprendente. Lo zoologo e anatomista francese Guillaume Rondelet (1507–1566) pubblicò tra gli altri anche importanti lavori di carattere ittiologico. In particolare, nella sua opera De piscibus marinis del 1554, denominò quello che oggi viene popolarmente indicato come “squalo martello” col nome di Libella marina per la somiglianza della testa di tale squalo con la “livella”, uno strumento medievale a forma di "T", impiegato dai falegnami. Sempre il Rondelet riprese questo nome nel 1555 per designare con Libella fluviatilis (= libella dei corsi d'acqua) la ninfa degli insetti che oggi chiamiamo Zigotteri; in effetti molte delle ninfe tra gli Zigotteri hanno una testa particolare, allargata e con gli occhi posti lateralmente in modo che, osservandone una nel suo insieme, essa può ricordare la forma di una "T". In seguito molti studiosi e naturalisti estesero l'uso del termine Libella a tutte le specie adulte (cioè anche alle imago e non solo alle ninfe) all’epoca conosciute. Sulla falsa riga di questa prassi nomenclaturale, anche Linneo (1707-1778) utilizzò il nome Libella ma scritto nel modo diminutivo (Libellula) nella prima edizione del suo Systema Naturae (1735). Non è dato sapere il motivo che indusse Linneo a coniare questo diminutivo: forse per significarne le piccole dimensioni o, più probabilmente, per seguire una prassi popolare che già indicava con vari diminutivi/vezzeggiativi alcune specie note all’epoca, come virguncula (da virgo) = piccola vergine o puellula (da puella) = fanciulla, signorina; o l’uso vernacolare francese dell’epoca, con Demoiselles (= signorine) per designare le specie più minute. Comunque sia, nella successiva decima edizione del Systema Naturae (1758), quella più famosa in cui appare per la prima volta la "nomenclatura scientifica binomiale", Linneo riprende il termine Libellula, ne fa un raggruppamento a sé e vi inserisce tutte le 18 specie all’epoca conosciute, descrivendole e classificandole tra i Neuroptera [dal greco νεῦρον (neuron) = nervo; e dal greco πτερόν (pteròn) = ala; in riferimento alle ali ricche di nervature]. Collocò poi queste specie in 2 gruppi: in un gruppo appaiono quelle descritte con Alis patentibus acquiescentes (= ali aperte a riposo) e corrispondenti a 16 specie degli attuali Anisotteri; nell'altro gruppo vi sono quelle descritte con Oculi distantes remotique (= occhi distanti e lontani) e corrispondenti a 2 degli attuali Zigotteri: Coenagrion puella e Calopteryx virgo. Da Linneo in poi, con il progredire dello studio e del riordinamento tassonomico e sistematico, il raggruppamento generale delle Libellula nel senso linneiano, prenderà il nome di Odonata e figurerà, tassonomicamente parlando, prima come sottordine e infine come ordine. Fu istituito nel 1793 dall’entomologo, botanico e medico danese Johann Christian Fabricius (1745–1808); l’etimologia, di cui il Fabricius non rilascia alcun riferimento scritto, potrebbe risalire al greco οδούς (odoùs), οδὀντος (odòntos) = dente, probabilmente in combinazione con una contrazione del termine greco γνάθος (gnàthos) = mascella; in riferimento al potente apparato masticatore di questi eccellenti cacciatori volanti. Altra possibile interpretazione etimologica, ma in definitiva con percorso simile, propone che il termine Odonata coniato dal Fabricius sia nato dalla contrazione del termine Odontognati, un raggruppamento di uccelli fossili risalenti al Cretaceo e caratterizzati dall’avere le mascelle provviste di denti. Il termine Libellula andrà nel seguito a designare (scientificamente parlando) solo un particolare genere di tale ordine, Libellula (Linnaeus, 1758), genere che oggi comprende diverse decine di specie ma che in Italia è rappresentato da 3 sole specie: Libellula depressa (Linnaeus, 1758), Libellula fulva (Müller, 1764) e Libellula quadrimaculata (Linnaeus, 1758). Osservazione: nell’attuale linguaggio corrente, comune e popolare, con il termine “libellule” vengono indicati indifferentemente tutti gli insetti appartenenti all’ordine Odonata, in ciò richiamando alla memoria quanto fu definito ai suoi tempi da Linneo. In un linguaggio altrettanto attuale e corrente ma più articolato, col termine “libellule” vengono indicate tutte le specie “più grandi e vistose” di tale ordine cioè, in pratica, le specie di insetti appartenenti al sottordine Anisoptera; questo per distinguerle dalle “damigelle” o “donzelle”, nomi popolari con cui vengono designate le specie appartenenti al sottordine Zygoptera. Infine, in un linguaggio propriamente scientifico, il termine Libellula indica esclusivamente il genere Libellula (Linnaeus, 1758). Nel seguito di questi appunti, all’occorrenza, potranno essere impiegati i termini “libellula” e/o “libellule” nelle tre suddette accezioni e il loro diverso significato sarà chiarito dal contesto. 2.2_ Caratteristiche essenziali degli insetti appartenenti all’ordine Odonata In alto: maschio immaturo di Trithemis annulata (Palisot de Beauvois, 1807) [sottordine Anisoptera; famiglia Libellulidae]; in basso: copula di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae] In alto: tandem in volo di Coenagrion scitulum (Rambur, 1842) [sottordine Zygoptera; famiglia Coenagrionidae]; in basso: femmina in ovideposizione di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae] Esempio di rappresentanti maschili dei due sottordini dell'ordine Odonata. A sinistra Calopteryx splendens (sottordine Zygoptera); a destra Onychogomphus forcipatus (sottordine Anisoptera). Pur avendo dimensioni simili in lunghezza (essendo i Calopterigidi tra gli Zigotteri più grandi e gli Onychogomphus tra gli Anisotteri più piccoli) la differenza che salta immediatamente agli occhi è il modo con cui vengono tenute le ali a riposo. Anche l'osservazione degli occhi non deve trarre in inganno: distanziati abbondantemente e relativamente piccoli tra gli Zigotteri; decisamente più grandi e o mai distanziati o un poco distanziati unicamente nelle specie della famiglia Gomphidae come, appunto, nel caso degli Onychogomphus. Le libellule sono insetti legati indissolubilmente agli ambienti acquatici, prevalentemente di acqua dolce (nei pressi di stagni, laghi, paludi, torbiere, pozze, ecc. o vari corsi d'acqua purché con correnti non troppo impetuose), ma pure con qualche specie reperibile presso acque più o meno salmastre come, ad esempio, Lestes macrostigma, Brachytron pratense, Aeshna mixta, Sympetrum fonscolombii e Ischnura genei. Presentano metamorfosi incompleta. Il loro ciclo biologico consiste, molto sinteticamente, nelle seguenti fasi: dall’uovo “fecondato e maturo”, depositato in ambiente acquatico o nelle strette vicinanze, nasce una neanide che quindi si trova già immersa in acqua o che comunque riesce a raggiungerla in breve tempo; la neanide si trasforma quasi subito in ninfa vera e propria che si accresce tramite un certo numero di mute, vivendo sempre in acqua; dopo l’ultima muta la ninfa esce dall’acqua e si trasforma (direttamente, cioè senza "impuparsi" come fanno ad esempio le farfalle) in insetto adulto (o imago), atto al volo e la cui vita trascorrerà tra terra e aria; passato qualche giorno (o poche settimane, a seconda della specie) gli adulti diventano sessualmente maturi e si accoppiano; parte delle uova della femmina viene fecondata dal maschio tramite copule dalla procedura del tutto particolare; quindi tutte le uova (fecondate o meno) vengono deposte nell’ambiente e cioè, a seconda della specie, o direttamente in acqua, o all’interno di steli di vegetazione acquatica sia sommersa che emersa, oppure in zone temporaneamente asciutte ma che, con la stagione delle piogge, torneranno ad essere sommerse; dopo un certo lasso di tempo variabile (in base alla specie) da poche settimane a qualche mese qualche uovo fecondato matura in embrione dal cui sviluppo nascerà la neanide e il ciclo si ripeterà. Dopo aver affrontato uno o solitamente più accoppiamenti, le libellule giungono alla fine del loro ciclo vitale e muoiono; in questo caso si tratta di “morte per vecchiaia”. La vita delle libellule adulte è relativamente corta e, escludendo le specie del genere Sympecma che svernano da adulte, può durare dalle 6 alle 8 settimane nelle specie più longeve (ad esempio in Ischnura elegans tra gli Zigotteri o Aeshna cyanea tra gli Anisotteri) ma più frequentemente viene consumata nell’arco di 2-3 settimane. Inoltre, pur rappresentando delle perfezionate “macchine volanti” atte alla caccia, anche le libellule incorrono in morti premature essendo esse stesse fonte di nutrimento per altri animali. Sono infatti un cibo agognato da anfibi e pesci ma anche da varie specie di uccelli che riescono a catturarle in volo; le si possono vedere (soprattutto le specie più piccole, in particolare, tra gli Zigotteri) cadute nelle trappole vischiose delle ragnatele e quindi destinate a morte sicura; e, infine, non è raro assistere alla scena in cui una libellula ha catturato e divorato una libellula più piccola ma talvolta anche di pari dimensioni, sia conspecifica che non. In Natura niente si "butta via"; tutto viene riciclato. Una femmina morta di Orthetrum coerulescens galleggia sull'acqua, una riserva di proteine per un gruppetto di Gerridi. Questo ragnetto ha costruito la sua trappola "ragnatelosa" tra la vegetazione a due metri dall'acqua. E ha funzionato benissimo, con alcune damigelle della specie Ischnura elegans che sono andate a impigliarvisi. Per le dimensioni del ragno occorre considerare che queste damigelle sono lunghe al massimo 3,5 cm. Un maschio di Anax parthenope ha catturato un maschio di Trithemis annulata e, portandolo in alto tra la vegetazione, si appresta a divorarlo. Foto di Massimo Squarcini. Un gruccione (Merops apiaster, Linnaeus, 1758) ha catturato un maschio di Orthetrum cancellatum. Foto di Gianni Bonini. Un gruccione (Merops apiaster, Linnaeus, 1758) ha catturato una femmina di Orthetrum albistylum. Foto di Gianni Bonini. Un airone rosso (Ardea purpurea, Linnaeus, 1766) ha appena catturato un maschio di Trithemis annulata. Foto di Gianni Bonini. Tuttavia se consideriamo anche il tempo trascorso nello stadio ninfale si può affermare che gli Odonati hanno, a seconda delle famiglie di appartenenza, una vita abbastanza lunga. Infatti alcune specie possono raggiungere i 4-5 anni di età allo stadio ninfale prima di affrontare l'ultima muta e il conseguente sfarfallamento (ad esempio alcune specie della famiglia Cordulegastridae); ma la loro vita allo stadio di imago, come accennato più sopra, si spenge nell'arco di poche settimane. Solo le specie del genere Sympecma hanno allo stadio di imago vita relativamente lunga perché solitamente sfarfallano in estate, riescono a svernare da adulte al riparo della vegetazione resistendo alle basse temperature, si accoppiano ai successivi primi tepori primaverili con conseguente deposizione delle uova, poi muoiono. Dalle loro uova nasceranno ninfe con stadio ninfale brevissimo visto che già dopo un paio di mesi affronteranno la metamorfosi in imago, ripetendo il ciclo. Le ninfe presentano, similmente a quanto si osserva negli adulti, 3 strutture principali: testa (munita di 2 occhi e di 2 antenne), torace (con 3 paia di zampe), addome. Negli stadi giovanili (brevissimo stadio di neanide e successivo più o meno lungo stadio ninfale, a seconda della specie) la loro vita trascorre in acqua. Gli Zigotteri respirano grazie a particolari e appariscenti pesudobranchie costituite da 3 grandi lamelle poste in coda all'addome; mentre gli Anisotteri respirano tramite numerose e minuscole lamelle pseudobranchiali disseminate nel tratto terminale (anale) dell’intestino. La loro alimentazione è esclusivamente carnivora essendo le ninfe ottime cacciatrici nei confronti di prede esclusivamente (tranne casi sporadici) viventi. Le specie più grandi possono catturare e divorare anche girini e addirittura avannotti. Ninfa di Zigottero: Calopteryx haemorrhoidalis (Vander Linden, 1825) [sottordine Zygoptera; famiglia Calopterygidae] Foto di Massimo Squarcini. Ninfe di Anisotteri: a sinistra Orthetrum brunneum (Fonscolombe, 1837); a destra un probabile Orthetrum coerulescens (Fabricius, 1798) [sottordine Anisoptera; famiglia Libellulidae]. Ninfa di Orthetrum brunneum (Fonscolombe, 1837) [sottordine Anisoptera; famiglia Libellulidae]; foto di Massimo Squarcini Tutte le ninfe, indipendentemente dalla specie, sono dotate di un organo particolare, perfettamente e implacabilmente adattato alla caccia, detto “maschera” ed esclusivo, in tutto il mondo animale, degli Odonati: infatti il loro labbro inferiore è modificato e sorregge due segmenti (mento e submento anch’essi modificati) allungati, articolati tra di loro e tenuti raccolti al di sotto della testa, con la sommità del segmento terminale che si pone davanti alla “faccia” (da cui nasce il nome “maschera"). Tale segmento terminale è munito all’apice di particolari uncini prensili che afferrano la preda quando la maschera, con una velocità e uno scatto sorprendenti (calcolabili in millesimi di secondo), viene estroflessa e allungata. Il paragone d’obbligo, tanto per rendere l’idea, viene fatto con l’arto superiore di un pugile che sta “in guardia”, cioè con l’arto che viene mantenuto raccolto (braccio e avambraccio stretti a contatto tra di loro e aderenti al corpo) tramite la piegatura del gomito e con il pugno a proteggere il volto (come una “maschera”); poi, “in attacco”, il gomito si apre e tutto l’arto scatta improvvisamente in avanti allungandosi e andando a colpire l’avversario anche a una certa distanza; la maschera delle ninfe funziona un po’ in tal modo, solo che, al posto del guantone del pugile, la maschera possiede delle appendici che afferrano la preda e poi la portano alla bocca. Difficilmente le ninfe, quando individuano la preda, sbagliano il colpo data la straordinaria velocità e precisione con cui effettuano l’attacco; per certi versi la funzionalità della maschera degli Odonati può ricordare l'analoga efficacia della lingua del camaleonte o delle zampe anteriori della mantide religiosa. Sopra: visione ventrale; sotto: visione laterale di una ninfa di Orthetrum brunneum (Fonscolombe, 1837) [sottordine Anisoptera; famiglia Libellulidae]; entrambe le foto sono di Massimo Squarcini Prendendo spunto da alcune foto di exuvia di Anax imperator visibili poco più sotto e pensando alla libellula ancora nello stadio ninfale, ecco (molto a grandi linee!) come si potrebbe presentare la maschera mentre sta scattando in avanti, in azione di caccia: Allo stadio ninfale le libellule si muovono più o meno lentamente sul fondo dell’ambiente acquatico in cui vivono: sia utilizzando le loro 3 paia di zampe per deambulare, sia muovendosi nuotando a scatti grazie all’emissione dalla estremità anale dell'addome di getti d’acqua, una sorta di "propulsione a reazione”. La ninfa subisce svariate mute di accrescimento, tra le 10-11 e le 14-15 mute a seconda della specie. Sempre in base alla specie e/o alle condizioni ambientali la vita ninfale degli Odonati può variare da pochi mesi a 4 o 5 anni. Con le ultime mute iniziano a svilupparsi particolari strutture tenute ben compresse e raccolte in apposite tasche (pteroteche) poste al di sopra del torace e che si trasformeranno successivamente in ali nell’insetto adulto. Al termine dell’ultima muta la ninfa esce dall’acqua riparandosi o arrampicandosi su qualche sostegno. Tale fase di emersione rappresenta uno dei momenti più delicati e vulnerabili nella vita di questi insetti poiché si trovano ad essere totalmente indifesi e alla mercé di potenziali predatori. Dalla ninfa così esposta all’aria emergerà infine, con varie metodologie e con tempi diversi a seconda della specie, l’insetto adulto che inizierà la respirazione aerea attraverso delle particolari piccole aperture (stigma) collocate sui fianchi del torace (gli stigma più grandi ed evidenti) e sull’addome. Un anisottero della famiglia Aeshnidae ha appena compiuto la metamorfosi, ed è ancora a contatto con la sua exuvia da cui è "emerso". Il suo corpo, in particolare l'addome non ha ancora raggiunto le dimensioni finali. Le raggiungerà pompandovi all'interno l'emolinfa. Anche le ali dovranno asciugarsi del tutto prima che possa affrontare il primo volo. Foto di Luigi Minciarelli. Femmina emergente di Lestes virens (Charpentier, 1825) [sottordine Zygoptera; famiglia Lestidae]; ha da poco compiuto l'ultima metamorfosi, "emergendo" dalla ninfa a cui rimane ancora momentaneamente vicina o attaccata, fino a quando il suo corpo, ali comprese, non si sarà asciugato in modo da permettergli di compiere il primo volo. Maschio emergente di Lestes virens (Charpentier, 1825) [sottordine Zygoptera; famiglia Lestidae]; ha da poco compiuto l'ultima metamorfosi, "emergendo" dalla ninfa a cui rimane ancora momentaneamente vicino o attaccato, fino a quando il suo corpo, ali comprese, non si sarà asciugato in modo da permettergli di compiere il primo volo. Femmine emergenti di Coenagrion scitulum (Rambur, 1842) [sottordine Zygoptera; famiglia Coenagrionidae]; hanno da poco compiuto l'ultima metamorfosi, "emergendo" dalla ninfa a cui rimangono ancora momentaneamente vicine o attaccate, fino a quando il loro corpo, ali comprese, non si sarà asciugato ed avrà acquisito le dimensioni finali, in modo da permettere di compiere il primo volo. Femmine emergenti di Coenagrion scitulum (Rambur, 1842) [sottordine Zygoptera; famiglia Coenagrionidae]; hanno da poco compiuto l'ultima metamorfosi, "emergendo" dalla ninfa a cui rimangono ancora momentaneamente vicine o attaccate. Nella foto qui sopra si vede chiaramente come l'esemplare di sinistra abbia l'addome non perfettamente disteso e allungato ma, al contrario, ancora corto e compresso; l'esemplare di destra ha già l'addome quasi del tutto allungato e disteso, mentre una gocciolina di emolinfa sta per essere espulsa dall'apertura anale. L'emolinfa viene "pompata" nell'addome e nelle ali proprio per permettere ai cosiddetti "tenerali" (altro nomignolo con cui vengono indicate le libellule appena emerse dalla ninfa) di raggiungere le dimensioni definitive; l'emolinfa in eccesso viene poi in qualche modo espulsa. In un certo lasso di tempo variabile da specie a specie ma solitamente entro poche ore, l’insetto espanderà l'addome pompandovi al suo interno sia l’emolinfa che l'acqua residua precedentemente accumulata; poi stenderà le ali sempre grazie all'emolinfa che vi viene immessa tramite le venature e al termine delle varie operazioni assumerà la forma finale e definitiva senza che le sue dimensioni crescano ulteriormente; viceversa e in generale sarà la sua colorazione che cambierà a seconda della età. Quando la libellula compirà il suo primo volo abbandonerà, sul supporto su cui si era ancorata la ninfa, una spoglia vuota (o exuvia) cioè l’involucro o esoscheletro della ninfa stessa. L’exuvia rappresenta quindi una sorta di “copia” della ninfa al suo stadio finale, una testimonianza prima dell’ultima e definitiva metamorfosi. Exuvie di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]; nella seconda foto è ben visibile la "maschera" raccolta in stato di riposo (più correttamente, si tratta dell'esoscheletro che la conteneva) e posta al di sotto della testa. Exuvia di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]; la freccia inferiore indica la maschera, a riposo, raccolta al di sotto della testa; la freccia superiore indica uno dei 2 uncini/artigli che si trovano all'apice della maschera, al culmine del palpo labiale. Maschio emergente di Sympetrum sanguineum (Müller, 1764) [sottordine Anisoptera; famiglia Libellulidae]; ha da poco compiuto l'ultima metamorfosi, "emergendo" dalla ninfa a cui rimane ancora momentaneamente vicino o attaccato, fino a quando il suo corpo, ali comprese, non si sarà asciugato in modo da permettergli di compiere il primo volo. Foto di Massimo Squarcini La libellula, fatto il primo volo, solitamente si ferma su qualche posatoio prossimo allo specchio d’acqua: sia per completare l’opera di asciugamento espellendo dall’addome l’emolinfa in eccesso sia per permettere la definitiva distensione e asciugatura delle ali; ali che, in questa prima fase di vita dell’imago, si presentano opalescenti. Maschio neosfarfallato di Orthetrum cancellatum (Linnaeus, 1758) [sottordine Anisoptera; famiglia Libellulidae]; notare le ali opalescenti, la traslucidità dell'addome e la gocciolina di emolinfa nella parte terminale dell'addome. Femmina neosfarfallata di Platycnemis pennipes (Pallas, 1771) [Sottordine Zygoptera; famiglia Platycnemididae]; notare le ali ancora opache e la trasparenza dell'addome entro cui si possono notare alcuni organi interni: il "vaso dorsale" (una sorta di cuore a sezioni contrattili) e il tubicino dell'apparato digerente (stomaco e intestino). Maschio neosfarfallato di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]; notare le ali opalescenti, i colori ancora molto neutri (rispetto a quelli che assumerà da adulto) e la gocciolina di emolinfa al di sotto del 2° segmento addominale. Raggiunto lo stadio adulto le libellule si riveleranno abili o abilissimi insetti volanti, esclusivamente carnivori ed eccezionali cacciatori prevalentemente di altri insetti volanti, tra cui talvolta non disdegnano di catturare e cibarsi di altre libellule di pari o solitamente più piccole dimensioni. Alcune specie vivono esclusivamente nei pressi dello specchio d’acqua di origine, altre amano allontanarsene anche di diverse centinaia di metri ma facendovi ritorno per la riproduzione; altre ancora possono affrontare migrazioni imponenti sia per numero di individui che per distanze percorse, anche di molte centinaia di chilometri. Alcuni generi di Odonati annoverano libellule che raggiungono taglie tra le più grandi nel mondo degli insetti. Per quanto riguarda gli Odonati europei e a seconda della specie e del sesso, l’apertura alare può variare da un minimo di 25-30 mm a un massimo di 110-115 mm; mentre la lunghezza totale del corpo può variare, sempre in base alla specie e al sesso, tra 25-30 mm e 80-90 mm. Maschio di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]; tra gli Odonati più grandi riscontrabili in Europa. Il suo corpo può raggiungere una lunghezza di 80-85 mm. Maschio di Anax ephippiger (Burmeister, 1839) [sottordine Anisoptera; famiglia Aeshnidae]; la sua apertura alare può raggiungere i 105-110 mm, tra le maggiori riscontrabili negli Odonati europei. Femmina di Ischnura pumilio (Charpentier, 1825) [sottordine Zygoptera; famiglia Coenagrionidae]; tra le libellule più minute reperibili in Italia, con lunghezza corporea minima sui 25 mm e apertura alare minima sui 33-35 mm. Le libellule adulte presentano un corpo allungato e più o meno snello in cui si individuano, ancor più facilmente che nelle ninfe, 3 particolari strutture: testa, torace e addome, con il torace che appare ben più robusto e tozzo rispetto all’addome. Dal torace spuntano tre paia di zampe praticamente mai impiegate per camminare sul suolo (contrariamente a quanto accadeva nella fase ninfale) ma, viceversa, utilizzate per la caccia in volo, per afferrare le prede o per appoggiarsi a vari posatoi, sui quali raramente le si vede fare brevi e pochi "passi" per meglio posizionarsi. Sempre dal torace si elevano le ali: due paia di ali direttamente collegate col torace, estremamente complesse nelle loro venature, molto grandi in proporzione al corpo, membranose e non ripiegabili e soprattutto di una stupefacente efficacia aerodinamica che fa di questi insetti delle vere e proprie meraviglie volanti. Le ali sono strettamente connesse a potenti muscoli situati all'interno del torace, muscoli che possono essere attivati indipendentemente l'uno dall'altro; ciò comporta che, all'occorrenza, anche le ali possono essere controllate e azionate indipendentemente le une dalle altre, permettendo alle libellule (soprattutto per le specie appartenenti al sottordine Anisoptera) di compiere qualsiasi evoluzione aerea, dalla planata allo stallo all'improvvisa virata, finanche alla “retromarcia”. In molti altri insetti, invece, il battito delle ali è sempre sincrono dipendendo dalla contemporanea contrazione e dal rilascio dell'intera muscolatura. La potente muscolatura che controlla le ali delle libellule consente a qualche specie di Anisottero di raggiungere scatti valutabili in velocità di circa 50 km/h mentre la frequenza dei battiti d’ala può sfiorare i 30 battiti al secondo. Calopteryx virgo (Linnaeus, 1758) [sottordine Zygoptera; famiglia Calopterygidae]; indipendenza delle ali di un maschio in volo. Aeshna mixta (Latreille, 1805) [sottordine Anisoptera; famiglia Aeshnidae]; indipendenza delle ali di un maschio in volo. Notare anche la disposizione delle zampe durante il volo. Negli Anisotteri le zampe vengono disposte nel modo più aereodinamico possibile per facilitare la velocità e l'agilità di volo: il paio di zampe anteriori è ripiegato e tenuto nascosto in verticale tra il retro degli occhi e il torace; la altre due paia di zampe sono tenute ripiegate e raccolte a contatto con la parte inferiore del torace. Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae]; indipendenza delle ali di un maschio in volo. Notare anche la disposizione delle zampe durante il volo. Negli Anisotteri le zampe vengono disposte nel modo più aereodinamico possibile per facilitare la velocità e l'agilità di volo: il paio di zampe anteriori è ripiegato e tenuto nascosto in verticale tra il retro degli occhi e il torace; la altre due paia di zampe sono tenute ripiegate e raccolte a contatto con la parte inferiore del torace. 2.3_ Testa, occhi e apparato masticatore Sulla testa fanno bella mostra di sé i due occhi, sia che si tratti di Anisotteri (con grandi occhi sempre più o meno posti a contatto tra di loro tranne che nella famiglia Gomphidae) o di Zigotteri (con occhi più piccoli, sempre nettamente separati tra di loro e posti ai lati della testa). Maschio di Aeshna affinis (Vander Linden, 1820) [sottordine Anisoptera; famiglia Aeshnidae]; grandi occhi che sono a contatto tra di loro per un lungo tratto. Maschio in volo di Anax ephippiger (Burmeister, 1839) [sottordine Anisoptera; famiglia Aeshnidae]; grandi occhi che sono a contatto tra di loro per un lungo tratto. Notare anche la disposizione delle zampe durante il volo. Negli Anisotteri le zampe vengono disposte nel modo più aereodinamico possibile per facilitare la velocità e l'agilità di volo: il paio di zampe anteriori è ripiegato e tenuto nascosto in verticale tra il retro degli occhi e il torace; la altre due paia di zampe sono tenute ripiegate e raccolte a contatto con la parte inferiore del torace. Maschio di Libellula depressa (Linnaeus, 1758) [sottordine Anisoptera; famiglia Libellulidae]; grandi occhi che sono a contatto tra di loro solo per un breve tratto. Maschio immaturo di Sympetrum fonscolombii (Sélys, 1840) [sottordine Anisoptera; famiglia Libellulidae]; grandi occhi che sono a contatto tra di loro solo per un breve tratto. Maschio di Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae]; grandi occhi che sono a contatto tra di loro solo per un breve tratto. Maschio di Cordulegaster boltonii (Donovan, 1807) [sottordine Anisoptera; famiglia Cordulegastridae]; grandi occhi che sono a contatto tra di loro in un solo punto. Maschio di Onychogomphus forcipatus subsp. unguiculatus (Linnaeus, 1758) [sottordine Anisoptera; famiglia Gomphidae]; grandi occhi nettamente separati tra di loro. Femmina di Platycnemis pennipes (Pallas, 1771) [sottordine Zygoptera; famiglia Platycnemididae]; occhi nettamente separati tra di loro, posti ai lati della testa che si presenta più larga che lunga. Maschio di Pyrrhosoma nymphula (Sulzer, 1776) [sottordine Zygoptera; famiglia Coenagrionidae]; occhi nettamente separati tra di loro, posti ai lati della testa che si presenta più larga che lunga. Maschio di Lestes virens (Charpentier, 1825) [sottordine Zygoptera; famiglia Lestidae]; occhi nettamente separati tra di loro, posti ai lati della testa che si presenta più larga che lunga. Maschio di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]; occhi nettamente separati tra di loro, posti ai lati della testa. Maschio di Coenagrion castellani (Roberts, 1948); [sottordine Zygoptera; famiglia Coenagrionidae]; occhi nettamente separati tra di loro, posti ai lati della testa. Maschio di Ischnura elegans (Vander Linden, 1820) [sottordine Zygoptera; famiglia Coenagrionidae]; occhi nettamente separati tra di loro, posti ai lati della testa. Femmina di Calopteryx haemorrhoidalis (Vander Linden, 1825); [sottordine Zygoptera; famiglia Calopterygidae]; occhi nettamente separati tra di loro, posti ai lati della testa. Maschio di Coenagrion castellani (Roberts, 1948) [sottordine Zygoptera; famiglia Coenagrionidae]; occhi nettamente separati tra di loro, posti ai lati della testa che si presenta più larga che lunga. Maschio di Calopteryx splendens (Vander Linden, 1825); [sottordine Zygoptera; famiglia Calopterygidae]; occhi nettamente separati tra di loro, posti ai lati della testa che si presenta più larga che lunga. Maschio di Erythromma lindenii (Sélys, 1840) [sottordine Zygoptera; famiglia Coenagrionidae]; occhi nettamente separati tra di loro, posti ai lati della testa che si presenta più larga che lunga. Maschio di Chalcolestes viridis (Vander Linden, 1825) [sottordine Zygoptera; famiglia Lestidae]; occhi nettamente separati tra di loro, posti ai lati della testa che si presenta più larga che lunga. Sono in ogni caso occhi composti da migliaia di piccoli e perfezionati elementi detti "ommatidi" e che, grazie anche all’estrema libertà di movimento che la testa può compiere muovendosi e ruotando in varie direzioni sul piccolo collo, consentono a questi insetti una visione panoramica ad ampissimo raggio. L’occhio composto, soprattutto degli Anisotteri, occupa una buona parte della testa tanto che alcune libellule tra le più grandi possono disporre fino a 20.000-25.000 ommatidi per ciascun occhio. In alcuni ingrandimenti si possono apprezzare anche le differenze di dimensione tra gli ommatidi: i più grandi sono collocati nella parte superiore dell’occhio e sono adatti per percepire gli oggetti in movimento (soprattutto le prede, ma anche gli eventuali predatori) e in lontananza, mentre gli ommatidi di sezione più piccola sono disposti nella zona inferiore dell’occhio e sono preposti alla visione ravvicinata e quindi ai dettagli o ai particolari. Gli ommatidi delle libellule percepiscono solo i raggi luminosi paralleli al loro asse longitudinale e in condizioni di buona luminosità, componendo poi a livello cerebrale l’immagine complessiva a mo’ di mosaico; l’esigenza della luminosità dei raggi incidenti giustifica il fatto che le libellule siano sostanzialmente insetti diurni con alcune eccezioni come, ad esempio, Boyeria irene o Anax ephippiger che possono essere attive anche nelle ore crepuscolari. Maschio immaturo di Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]; i suoi occhi sono composti da migliaia di ommatidi, qui visibili solo quelli delle zone superiori. Femmina immatura di Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae]; migliaia di ommatidi compongono i suoi occhi; nella foto è anche evidente, grazie alla diversa colorazione naturale, la separazione tra le parti superiore (marroncina, con gli ommatidi di sezione maggiore) e inferiore (verde-giallognola, con ommatidi di sezione minore). Maschio di Orthetrum coerulescens (Fabricius, 1798) [sottordine Anisoptera; famiglia Libellulidae]; migliaia di ommatidi compongono i suoi occhi. Abbastanza evidenti quelli della parte superiore dell’occhio; non ben distinguibili quelli della parte inferiore perché di sezione minore. Maschio immaturo di Crocothemis erythraea (Brullé, 1832) [sottordine Anisoptera; famiglia Libellulidae]; migliaia di ommatidi compongono la zona superiore (marroncina-rosata) dei suoi occhi; nella zona inferiore (celestina) gli ommatidi sono di sezione più piccola. Maschio di Anax imperator (Leach, 1815) [sottordine Anisoptera; famiglia Aeshnidae]; migliaia di ommatidi compongono la zona superiore (azzurra) dei suoi occhi; nella zona inferiore (verdolina) gli ommatidi sono di sezione più piccola. Maschio immaturo di Orthetrum albistylum (Sélys, 1848) [sottordine Anisoptera; famiglia Libellulidae]; migliaia di ommatidi compongono la zona superiore (rosata) dei suoi occhi; nella zona inferiore (marroncina) gli ommatidi sono di sezione più piccola. Femmina di Trithemis annulata (Palisot de Beauvois, 1807) [sottordine Anisoptera; famiglia Libellulidae]; parte superiore (bruno-rossiccia) degli occhi, con gli ommatidi più evidenti. Maschio immaturo di Sympetrum sanguineum (Müller, 1764) [sottordine Anisoptera; famiglia Libellulidae]; migliaia di ommatidi compongono la zona superiore (rosata) dei suoi occhi; nella zona inferiore (marroncina) gli ommatidi sono di sezione più piccola. Foto di Massimo Squarcini Femmina immatura di Aeshna affinis (Vander Linden, 1820) [sottordine Anisoptera; famiglia Aeshnidae]; migliaia di ommatidi compongono la zona superiore (azzurra) dei suoi occhi; nella zona inferiore (verdolina) gli ommatidi sono di sezione più piccola. L’estrema funzionalità dell’articolazione tra il piccolo e (apparentemente) esile collo e la testa consente alle libellule di muovere la testa stessa sia lateralmente che in direzione alto-basso, ma anche in una straordinaria rotazione attorno al collo. Questa “abilità rotazionale” è facilmente verificabile osservando le libellule, grandi o piccole che siano, quando compiono una delle funzioni fondamentali per la loro “igiene”: la pulizia degli occhi. Con le zampe anteriori impiegate a mo’ di tergicristallo riescono a detergere gli occhi dalle varie impurità che vi si possono depositare sopra; la superficie oculare che riescono a raggiungere con le zampe anteriori è pressoché completa grazie proprio alla possibilità di ruotare la testa attorno al collo, anche di 180°. Sympetrum fonscolombii (Sélys, 1840) [sottordine Anisoptera; famiglia Libellulidae]; femmina che si sta detergendo gli occhi. L’operazione viene effettuata con le zampe anteriori che, a mo’ di un tergicristallo, tolgono dagli occhi le varie impurità accumulate. Oltre alla consueta separazione visibile nell’occhio tra parte superiore (brunastra, contenente gli ommatidi più grandi) e parte inferiore (celestina, contenente gli ommatidi più piccoli) è evidente la capacità di ruotare la testa attorno all’esile collo; nel caso della prima delle 3 foto precedenti, una rotazione di quasi 180° è stata necessaria per arrivare a detergere una maggior superficie oculare. Aeshna cyanea (Müller, 1764) [sottordine Anisoptera; famiglia Aeshnidae]; maschio che pulisce gli occhi. Come nel caso della foto precedente l’operazione viene effettuata con le zampe anteriori che, a mo’ di un tergicristallo, tolgono dagli occhi le varie impurità accumulate. Qui la testa è stata facilmente ruotata di circa 120°; visibile anche il non indifferente apparato masticatore con una sorta di denti aguzzi. Sympetrum fonscolombii (Sélys, 1840) [sottordine Anisoptera; famiglia Libellulidae]; maschio e pulizia degli occhi. Qui la testa è stata facilmente ruotata di 90°; degli occhi è visibile solo la parte inferiore (azzurrina), quella contenente gli ommatidi di sezione minore. Calopteryx haemorrhoidalis (Vander Linden, 1825) [sottordine Zygoptera; famiglia Calopterygidae]; femmina che si pulisce l’occhio destro dopo aver ruotato la testa. Coenagrion scitulum (Rambur, 1842) [sottordine Zygoptera; famiglia Coenagrionidae]; maschio che pulisce gli occhi. Questo maschio, nonostante la mancanza della zampa anteriore destra e del tarso della zampa anteriore sinistra, riesce comunque a detergersi gli occhi grazie alla facilità con cui può ruotare la testa attorno al collo. Maschio di Erythromma lindenii (Sélys, 1840) [sottordine Zygoptera; famiglia Coenagrionidae]; la zampa anteriore destra, grazie alla rotazione della testa attorno al collo, può detergere una buona parte dell’occhio destro. Femmina di Coenagrion castellani (Roberts, 1948) [sottordine Zygoptera; famiglia Coenagrionidae]; impegnata con la pulizia degli occhi Maschio immaturo di Trithemis annulata (Palisot de Beauvois, 1807) [sottordine Anisoptera; famiglia Libellulidae] che si deterge gli occhi; con rotazione della testa di 90° Maschio di Lestes barbarus (Fabricius, 1798) [sottordine Zygoptera; famiglia Lestidae]; si pulisce gli occhi e gli ocelli, grazie a una rotazione della testa di oltre 90°. Femmina di Sympetrum striolatum (Charpentier, 1840) [sottordine Anisoptera; famiglia Libellulidae] che si deterge gli occhi; nell'ultima foto: con rotazione della testa di quasi 180°

.jpg.2e622a215b1e6e3dfe091d81aa1e8dde.jpg)

.jpg.50a96c348f66440871b9d963a2455942.jpg)
.jpg.60c7d60313657d9ecaceaa2e5c669533.jpg)
.jpg.48268dd190517d220264e96cb01c5bdd.jpg)

.jpg.3d31ec17ca866c0d3e9beb4d41cad3cd.jpg)
.jpg.84665b992ccf421c1a876c7f0925ec17.jpg)
.jpg.3a585f852527f446818868902893b2af.jpg)
.jpg.488e1441c079e6527c53d968a3b0b2ff.jpg)
.jpg.402c671ec61fd00e18f221b60d8a5f65.jpg)
.jpg.320c3fb2aee2ed7af1623040affcb69b.jpg)


.jpg.8ee1519272abe772fd47599683b527a1.jpg)
.jpg.981e67d42b73f77ee2449191953813da.jpg)
.jpg.625a41cf1b0d11317745a997b0542486.jpg)





















.jpg.716d5d99d3ac76c1475135369c050345.jpg)
.jpg.519ea2e56e7e798d67d6ba8ec13a9a81.jpg)
.jpg.a07bec88113af4a7d7e93271a11e4662.jpg)
.jpg.7231cb828173e941d32ff8fb14f32a95.jpg)
.jpg.5871b9c6b18d3ead919a2f7c75dd5a5d.jpg)
.jpg.c23998b27017b75b76954f44a91818b5.jpg)








---Copia.jpg.30cddd32a8880e5a809bf21abd8be0d0.jpg)





.jpg.e3367b3f9dae1a08f4b7beaf9ef075fa.jpg)
.jpg.52b80dc5cf709616c6d9d20926470e50.jpg)




.jpg.bcceae8389b510642126c7ca1208967d.jpg)
.jpg.38bf7db71cefb5ea14a9599f21973585.jpg)
.jpg.c455b9ab75493a89ab3801dafea690a7.jpg)
.jpg.c177197f3e8efd3e37c762bce0f0535c.jpg)
.jpg.790703e3a80d7e002e2939201ccc0cd0.jpg)




.jpg.68df187ed380e5bd617c30cb81154dcc.jpg)




-
ODONATA_ 01 - Il “percorso classico” tassonomico che conduce all’ordine Odonata
Alessandro F ha postato un topic nell'area Odonata
Il “percorso classico” tassonomico che conduce all’ordine Odonata Dominio: Eukaryota; è, assieme al dominio Prokariota, uno dei due domini della "classica" scuola tassonomica con cui vengono classificati gli organismi viventi uni- o pluricellulari. Dal greco εὖ (eu) = buono, e κάρυον (kàryon) = nucleo; in riferimento alla presenza, all'interno delle singole cellule di tali organismi, di un nucleo ben definito e separato tramite una o più membrane dal resto della cellula stessa. Il nucleo assume una importanza rilevante perché contiene la maggior parte del materiale genetico. Nel dominio Eukaryota troviamo, tra gli altri, il regno Animalia (quello che ci interessa da vicino), il regno Fungi (quello dei funghi) e il regno Plantae (quello delle piante). Il dominio Prokariota, dal greco πρό (pro) = prima, comprende organismi viventi unicellulari microscopici, in cui le cellule sono prive di un nucleo ben strutturato; ad esempio i batteri. Regno: Animalia; parola che deriva dal latino animalis = dotato di vita, di anima; a sua volta derivante dal greco άνεμος (ànemos) = vento, soffio. Quindi nel senso onnicomprensivo di "essere vivente". Veramente innumerevoli i phyla che costituiscono il regno Animalia; solo per fare alcuni esempi: Chordata (muniti di una sorta di colonna o "corda" dorsale, come i vertebrati), Anellida (gli anellidi, come i lombrichi), Mollusca (i molluschi, come le seppie o i calamari), Porifera (come le spugne), ecc. Tra i vari phyla quello che ci interessa è il seguente. Phylum: Arthropoda; dal greco ἄρθρον (àrthron) = articolazione, e ποδός (podòs) = piedi; quindi, alla lettera, “con i piedi articolati”; cioè con le zampe munite di articolazioni. Gli artropodi sono inoltre caratterizzati dal possedere un esoscheletro composto da chitina, un materiale leggero ed elastico ma anche resistente e duro a tal punto da proteggere e sostenere il corpo che racchiude. L'esoscheletro, pur nella sua elasticità, non è comunque in grado di aumentare la propria superficie durante l'accrescimento in età e dimensioni del corpo che racchiude. La crescita dell'animale avviene quindi grazie ad un susseguirsi di varie mute; ad ogni nuova muta l'organismo abbandona il vecchio esoscheletro (che "gli stava stretto") si espande in dimensioni quanto occorre (rimanendo momentaneamente "nudo") e si costruisce un nuovo esoscheletro più adatto alle nuove dimensioni acquisite. Gli artropodi sono muniti di un efficiente apparato respiratorio, sia che si tratti di organismi acquatici che terrestri. Sono per la maggior parte ovipari, cioè si riproducono tramite uova rilasciate all'esterno del corpo "materno" e il cui sviluppo embrionale avviene necessariamente senza scambio di materiale nutritivo con il progenitore; in alcuni casi sono ovovivipari (come, ad esempio, gli scorpioni) in cui le uova si sviluppano sì all’interno del corpo materno ma comunque senza scambio di nutrienti. Le uova prodotte dalla femmina, soprattutto se ovipara, sono solitamente numerosissime ma non tutte arriveranno ad essere fecondate e tra queste solo poche riusciranno a trasformarsi in embrioni e a dar vita a nuovi esemplari. Quello degli Arthropoda è il phylum più ricco di specie di tutto il regno animale; in effetti, a loro volta, gli Arthropoda sono suddivisibili in numerosi subphyla e superclassi come, ad esempio: subphylum Crustacea (i vari crostacei: gamberi, aragoste, granchi, ecc.); superclasse Myriapoda (come i millepiedi e simili); subphylum Chelicerata (come i ragni, i limuli o gli scorpioni), superclasse Hexapoda (quella che, in questo percorso, ci interessa). Superclasse: Hexapoda; dal greco ἑξα (èxa) = sei, e ποδός (podòs) = piedi; quindi, alla lettera: “con sei piedi”; in riferimento alla presenza di sei zampe, disposte in 3 paia. Le classi in cui si suddividono gli Hexapoda sono: Entognatha e, quella che qui ci interessa, Insecta. Classe: Insecta; dal latino insectus, che è participio passato del verbo latino insecare = tagliare, suddividere; in riferimento alla suddivisione del corpo in parti morfologicamente ben distinguibili; sostanzialmente e a grandi linee: testa, torace e addome. La classe Insecta (Linnaeus, 1758) è la più vasta classe, in numero di specie, di organismi viventi racchiudendone ben oltre un milione tra quelle attualmente classificate oltre a quelle (presumibilmente altrettante se non di più) ancora sconosciute. Si suddivide in due sottoclassi: la sottoclasse Apterygota (che annovera piccoli insetti privi di ali e che crescono in dimensioni solo attraverso le mute ma senza alcun tipo di metamorfosi mancando i vari stadi di ninfa e di pupa; quindi il loro stadio adulto è morfologicamente uguale allo stadio giovanile ma solo più grande) e la sottoclasse che riguarda il nostro percorso, la Pterygota. Sottoclasse: Pterygota; dal greco πτερυγωτός (pterugotòs) = alato; dal greco πτερόν (pteròn) = ala; in riferimento alla presenza di ali, solitamente in numero di 2 (un paio) o di 4 (due paia). A loro volta gli Pterygota vengono suddivisi in due coorti: Endopterygota e Exopterygota, di cui può essere utile sottolineare alcune caratteristiche differenziali. Coorte: Endopterygota; dal greco ἔνδον (èndon) = dentro, internamente. Annovera gli insetti alati più evoluti e che presentano metamorfosi completa. A grandi linee: dall’uovo fecondato e dallo sviluppo del conseguente embrione nasce la cosiddetta larva; segue poi lo “stadio di pupa” e le ali iniziano a formarsi proprio durante questo stadio in particolari strutture interne alla pupa stessa; dalla pupa "nascerà" l'insetto adulto (o imago); è il caso, ad esempio delle farfalle (in cui la pupa è anche detta crisalide), delle api o delle mosche. Coorte: Exopterygota; dal greco ἔξω (èxo) = fuori, esternamente. Annovera gli insetti alati più primitivi e che presentano metamorfosi incompleta. A grandi linee: dall’uovo fecondato e dallo sviluppo del conseguente embrione nasce una "neanide" trasformata quasi immediatamente nella cosiddetta ninfa che, grazie alle varie mute, crescerà in dimensioni. Le ali iniziano ad apparire negli ultimi “stadi di ninfa”; si tratta comunque di ali non funzionanti ma solo abbozzate e che si vanno a formare in particolari strutture o teche esterne alla ninfa stessa. La metamorfosi è incompleta perché non passa attraverso lo "stadio di pupa”. L'insetto adulto "nascerà" direttamente dalla ninfa quando questa avrà completato l'ultima sua muta. È il caso che ci interessa, quello in cui saranno inserite, tra gli altri, anche le libellule. Gli Exopterygota sono a loro volta suddivisi in due sottocoorti: Neoptera e Palaeoptera e di cui può essere utile rilevare le diversità. Sottocoorte: Neoptera; dal greco νέος (nèos) = nuovo. Le ali degli insetti adulti non sono direttamente collegate al torace, in quanto tra le ali e il torace vi sono delle brevi articolazioni più o meno elasticamente snodabili che permettono a questi insetti, a riposo, di ripiegare le ali “ad astuccio” e riporle a diretto contatto sull’addome in modo da essere maggiormente protette. È il caso, ad esempio, delle mantidi, dei grilli, delle cavallette, delle blatte, ecc. Sottocoorte: Palaeoptera; dal greco παλαιός (palaiòs) = antico. Negli insetti adulti le piccole articolazioni presenti tra torace e ali non sono né snodabili né così efficaci come accade tra i Neoptera. Tale peculiarità obbliga questi insetti, quando sono a riposo, a tenere le ali sempre spiegate in tutta la loro ampiezza; quindi non ripiegate né particolarmente protette. A sua volta la sottocoorte Palaeoptera è suddivisa in due ordini: l'ordine Ephemeroptera e, finalmente, quello che ci interessa da vicino, quello delle libellule in senso lato: ordine Odonata. Tra gli Ephemeroptera troviamo le cosiddette “effimere”, nome che deriva dal greco ἐϕήμερος (efìmeros) = effimero; a sua volta composto dal greco ἐπί (epì) = sopra e ἡμέρα (imèra) = giorno; cioè che dura solo un giorno, di breve durata. Si tratta di piccoli insetti con stadio ninfale in acqua dolce e stadio adulto terrestre; di vita assai breve nella fase di imago, sono mediocri volatori anche a causa delle dimensioni ridottissime delle ali posteriori che possono quasi mancare del tutto. Una "effimera", Cloeon dipterum (Linnaeus, 1761). Ordine Ephemeroptera; si tratta di una specie che, dopo le "schiuse" e conseguenti sciamature, ama rifugiarsi in ambienti ben protetti come nelle abitazioni in vicinanza a corsi d'acqua (non inquinati); è quindi facile osservare questo insetto su varie superfici casalinghe: dai muri ai vetri, sulle tende o sui mobili. Altra "effimera", in volo; stavolta del genere Ecdyonurus Eaton, 1868 (ordine Ephemeroptera). Una "libellula", Ischnura elegans (Vander Linden, 1820). Ordine Odonata. Una "libellula", Sympetrum striolatum (Charpentier, 1840). Ordine Odonata. Ordine: Odonata (Fabricius, 1793); gli insetti di questo ordine sono esclusivamente carnivori sia allo stadio ninfale che allo stadio adulto. Sono legati all’ambiente acquatico in quanto lo stadio ninfale si sviluppa completamente in tale habitat mentre le imago hanno vita terrestre e aerea. Sono insetti tipicamente ovipari. Le ninfe, unico caso nel mondo animale, sono dotate di un particolare organo raptatorio, detto "maschera", e posizionato sotto alla testa. Gli adulti sono facilmente riconoscibili tra tutti gli altri insetti grazie alle seguenti peculiarità: due grandi occhi composti e, viceversa, due piccole antenne; un addome assai lungo rispetto al resto del corpo; sono abili o abilissimi cacciatori volanti grazie a 2 paia di grandi ali ricche di venature e particolarmente specializzate e adatte al volo; sono inoltre dotati di un efficace apparato masticatore; alcune specie si collocano tra gli insetti più grandi e spesso anche tra i più colorati e variopinti. I maschi presentano due diversi organi atti alla riproduzione: uno sbocco seminale (che contiene lo sperma) nella parte posteriore dell’addome e un apparato meccanico-copulatore (una sorta di pene) nella parte anteriore dell’addome; questa particolare conformazione comporta una modalità del tutto esclusiva nella fase di accoppiamento. Gli Odonati sono presenti in ogni regione terrestre (habitat permettendo), ad esclusione dell’Antartide e di qualche isola artica. La sistematica “tradizionale” suddivide l’ordine Odonata nei seguenti 3 sottordini: Zygoptera (Sélys, 1854), Anisoptera (Sélys, 1854) e Anisozygoptera (Handlirsch, 1906). Una suddivisione più moderna, susseguente a vari studi effettuati alla fine del 20° secolo e confermati da successive analisi filogenetiche, prevede invece la suddivisione dell’ordine Odonata in soli 2 sottordini: Zygoptera (Sélys, 1854) e Epiprocta (Lohmann, 1996) con quest’ultimo a sua volta suddiviso negli infraordini Epiophlebioptera (Lohmann, 1996) (corrispondente al “tradizionale” e precedente Anisozygoptera, qui “declassato” a infraordine) e Anisoptera (Sélys, 1854) (anch’esso qui “declassato” a infraordine). Qualunque sia la scelta e rimanendo in un ambito europeo i taxa che riguardano gli Odonati sono esclusivamente Zygoptera e Anisoptera visto che gli Anisozygoptera (o Epiophlebioptera che dir si voglia) annoverano pochissime specie reperibili solo nel continente asiatico: 4 specie soltanto, secondo la World Odonata List, cioè una specie giapponese, una himalayana e due cinesi, tutte appartenenti al genere Epiophlebia (Calvert, 1903). Nel seguito di questi e di altri appunti verrà mantenuta la suddivisione “tradizionale” (come, del resto, viene tuttora accettato dalla World Odonata List) e quindi Anisoptera figurerà sempre come sottordine, al pari di Zygoptera. Secondo recenti dati della World Odonata List (Agosto 2023), a livello mondiale le specie riconosciute di Odonati sono poco più di 6400, inserite in quasi 700 generi, a loro volta raggruppati in una cinquantina di famiglie. A livello europeo le specie riconosciute sono circa 170, inserite in 47 generi, a loro volta inseriti in 11 famiglie. Mentre nel territorio italiano sono presenti (a meno di migrazioni, di periodiche assenze o di probabili estinzioni) 96 specie (o 95 secondo alcune fonti), inserite in 38 generi (12 tra gli Zigotteri e 26 tra gli Anisotteri), a loro volta facenti parte di 10 famiglie: Calopterygidae, Coenagrionidae, Lestidae e Platycnemididae tra gli Zigotteri; Aeshnidae, Cordulegastridae, Corduliidae, Gomphidae, Libellulidae e Synthemistidae tra gli Anisotteri.

.jpg.b4f5d1179ac9ea4c09b7403ea77a04f5.jpg)



-
Spiranthes spiralis (L.) Chevall.
Alessandro F ha postato un topic nell'area Orchidee Toscana Anno 2023
Spiranthes spiralis (L.) Chevall. (1827); Regione Toscana (Mugello; "Macchie di Panna"; 500 m s.l.m.); 28 Settembre 2023; Foto di Alessandro Francolini. Le ultime Orchidee a fiorire in Italia.




-
Orchis anthropophora (L.) All.
Alessandro F ha risposto alla discussione di Pietro Curti in Orchidee Umbria Anno 2023
Una delizia Ale -
Anacamptis pyramidalis (L.) Rich.
Alessandro F ha risposto alla discussione di Pietro Curti in Orchidee Umbria Anno 2023
Belle belle Un soffio quasi primaverile Ale -
Nigritella rhellicani Teppner & E.Klein
Alessandro F ha postato un topic nell'area Orchidee Trentino Alto Adige Anno 2023
Nigritella rhellicani Teppner & E.Klein; Regione Trentino Alto Adige, Val d'Ultimo (BZ); 2100 m s.l.m.; 14 Luglio 2023; su praterie ancora non adibite a pascolo bovino; foto di Alessandro Francolini. Con profumo molto gradevole, a seconda del fiore (o del vento?) variabile dalla vaniglia al cioccolato.








-
Gymnadenia conopsea (L.) R. Br. (1813)
Alessandro F ha risposto alla discussione di Alessandro F in Orchidee Trentino Alto Adige Anno 2023
Grazie ragazzi Ale -
Platanthera bifolia var. subalpina Brügger 1886
Alessandro F ha risposto alla discussione di Alessandro F in Orchidee Trentino Alto Adige Anno 2023
Grazie ragazzi Mi sono dimenticato di metterci la data ; scusatemi . Non siamo in agosto; le foto risalgono al 14 luglio 2023. Ale -
Platanthera bifolia var. subalpina Brügger 1886
Alessandro F ha postato un topic nell'area Orchidee Trentino Alto Adige Anno 2023
Platanthera bifolia var. subalpina Brügger 1886; Regione Trentino Alto Adige, Val d'Ultimo (BZ); 2100 m s.l.m. su praterie ancora non adibite a pascolo bovino; foto di Alessandro Francolini. Unica piantina avvistata (e ormai un po' sofferente), in mezzo a un mare di Gymnadenia conopsea.


-
Gymnadenia conopsea (L.) R. Br. (1813)
Alessandro F ha postato un topic nell'area Orchidee Trentino Alto Adige Anno 2023
Gymnadenia conopsea (L.) R. Br. (1813); Regione Trentino Alto Adige, Val d'Ultimo (BZ); 2100 m s.l.m.; 14 Luglio 2023; su praterie ancora non adibite a pascolo bovino; foto di Alessandro Francolini.






-
Epipactis palustris (L.) Crantz 1769.
Alessandro F ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2023
Favolose Ale -
Dactylorhiza lapponica subsp. rhaetica H. Baumann & R. Lorenz 2005
Alessandro F ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2023
Bella bella Ale -
Anacamptis pyramidalis (L.) Rich.
Alessandro F ha risposto alla discussione di Mario Iannotti in Orchidee Umbria Anno 2023
Bellissime Piramidi Ale -
Neottia cordata (L.) Rich.
Alessandro F ha risposto alla discussione di Giuliano Gnata in Orchidee Trentino Alto Adige Anno 2023
Macché mesto Sono molto belle lo stesso Ale -
Epipactis palustris (L.) Crantz 1769.
Alessandro F ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2023
Bellissima Tra le mie preferite, anche se non l'ho mai vista dal vivo Ale -
Nigritella nigra subsp. rhellicani (Teppner & E. Klein) H. Bauman, Kùnkele & L. Lorenz.
Alessandro F ha risposto alla discussione di Giuliano Gnata in Orchidee Trentino Alto Adige Anno 2023
Pienamente d'accordo Un vero spettacolo di forme e colori Da perderci la testa Ale -
Neotinea tridentata (Scop.) R.M. Bateman Pridgeon & M.W. Chase
Alessandro F ha risposto alla discussione di Pietro Curti in Orchidee Marche Anno 2023
Esuberanti Ale -
Nigritella nigra subsp. austriaca (Teppner & E. Klein) P. Delforge.
Alessandro F ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2023
Perfezione in miniatura Ale
.jpg.059a9c323114c8f0dad73b778c569f72.jpg)